


All Issues
Ecological research: Long-term studies to gauge effects of invading bees
Publication Information
California Agriculture 46(1):20-23.
Published January 01, 1992
PDF | Citation | Permissions
Abstract
The expected invasion of the United States by Africanized honey bees has inspired long-term studies documenting the ecological importance of native and introduced bees. Baseline data are being gathered to predict the effects of the invasion. Standardized sampling procedures and tools have been developed to monitor bee communities. The studies will provide information for developing wildland area conservation policies.
Full text

Above the sunflower bee (Diadasia enavata), a solitary bee, collects pollen at UC Davis farm. At left, the honey bee (Apis mellifera) robs pollen from the sunflower bee. Studies showed that host-specific native sunflower bees show no decrease in frequency near versus far away from an apiary, while honey bees and Megachile apicalis frequencies are inversely related.

The imminent invasion of the United States by Africanized honey bees has led to a long-term study of the diversity, floral resource use, and ecological interactions of native and introduced bees in wildlands of Central California. The study, initiated in 1987, will furnish baseline data on community structure, temporal patterns, and resource overlap of resident bees to prepare for the disruptions likely after the invasion. In addition, it will provide data on the ecology of invasions by other non-native bees. Insights gained from this study are expected to shape conservation policies.
The study will also supply ecological information on solitary bees, their role as pollinators of native plant species, and their potential use in crop pollination. The value of alternative pollinators will increase if production of sufficient honey bees for crop pollination is impaired by new mite parasites and the Africanized honey bee.
Honey bees and native wildlife
The honey bee, Apis mellifera L., is not native to the New World or Australasia. It has become naturalized and is so common in these areas that few realize it is an introduced species. The honey bee is not only vital to agriculture, especially in its role in crop pollination, but it also pollinates many wildflower species, enhancing their reproduction and providing fruits and seeds as forage for wildlife.

Oak tree with sampling units for monitoring bee nesting activities. Inset shows block sampling unit (BSU), 1990 design, for cavity-nesting solitary bees. Below is swarm trap for feral honey bees.
While honey bees have potential value in restoration projects, where native pollinators may have been eliminated, the full impact of this alien species on our wild-land and conserved areas is not well understood. It has been suggested that honey bees reduce populations of native pollinators, through competition for food, and that they interfere with reproduction of native plants, especially in Australia where much of the flora is adapted for pollination by birds. They may also enhance reproduction of weeds, such as yellow star thistle in California, and occupy cavities that could otherwise be used by nesting birds and mammals. More concise determinations of benefits and losses due to honey bees can assist in making sound policies regarding management of conserved areas and public wildlands.
Africanized honey bees
It is timely to determine the impact of honey bees on today's environment and to gather baseline ecological data on European honey bees (EHB) in North America before Africanized honey bees (AHB) arrive. AHB have displaced or hybridized with feral EHB throughout most of the New World tropics. Differences in colony defense, population size, growth rates, swarming, foraging, pollen and nectar hoarding, and colony densities suggest that feral populations of AHB may differ in ecological relationships from EHB in our wildlands.
AHB regional project
Scientists in American universities and in the U.S. Department of Agriculture have been attempting to understand this tropical race of honey bees and its phenomenal spread in the Americas so that its impact on crop pollination, beekeeping, and public health in the U.S. can be mitigated. Changes in beekeeping required with the expected presence of AHB are not compatible with pollination services, the most important economic use of honey bees in California (Gary et al., California Agriculture Nov-Dec 1985, pp. 4-7). In the fall of 1988 a national cooperative research effort was launched to establish Regional Project W-180, “Identification, behavioral ecology, genetics and management of African honey bees.” The projecf s 50 scientists from 15 states include 9 Agricultural Experiment Station universities, 5 USDA bee laboratories, and 4 other university scientists.
Our interest in bee communities focuses on three of W-180's six objectives: (1) to determine population densities of Apis and non-Apis bees before and after invasion of AHB; (2) to determine factors in the nest environment that influence population ecology and life histories of bees; and (3) to evaluate the floral resource carrying capacity of bees in order to predict effects of AHB. We have been gathering information on the diversity, phenology, relative abundance, and associations of flowers and bees in Central California's wildlands. These data are being used to determine patterns of food resource use, abundance and reproductive success of bees to detect potential competition between honey bees and solitary bees.
Study techniques
Six sites in the San Joaquin Valley and adjacent foothills were chosen to represent diverse wildland areas where AHB may survive (fig. 1). Valley sites include Cosumnes River (CO) and Creighton Ranch (CR) preserves (The Nature Conservancy), and San Luis National Wildlife Refuge (SL) (U.S. Fish and Wildlife Service). Foothill sites include Hastings Natural History Reservation (HR) (University of California Natural Reserve System), San Joaquin Experimental Range (SJ) (California State University at Fresno and U.S. Forest Service), and Mount Diablo State Park (California Department of Parks and Recreation). (Data from the last site are not yet available.)
During periods of adult bee activity we collect and observe bees, adding to data bases on diversity and relative abundance of bees and flowers, stages of floral bloom and floral associations (all flowers visited, not just those preferred by bees).
Selected flowering species, shared as food resources by honey bees and solitary bees, are monitored to determine potential competition for food. For example, we measured activity on Helianthus (sunflower) and Grindelia (gumweed) at SL in 1989, collecting timed samples and making nondestructive visual counts of bees near and away from an apiary. We have yearly repeated visual counts since 1989.
Nesting biologies, seasonality, relative abundance, population ecology, and site and hole sizes chosen by selected species of cavity-nesting solitary bees are assessed with block-sampling units (BSU) containing sticks with drilled holes. Based on preliminary data gathered since 1987, we standardized our BSU, numbers of BSU per site, and sampling frequency and seasons in 1990. Our standard BSU now consists of a block of 12 sticks: each stick is 19 by 23 by 117 mm with a hole 100 mm deep, and with four sticks each containing holes 4.7, 6.3 or 7.8 mm in diameter. The nine BSUs per site are replaced every 3 weeks from the end of February to the end of May at the foothill sites and from the end of May to early October at the valley sites. Holes are visually inspected initially to determine occupancy.
Most occupied sticks are X-rayed to provide more detail on the kinds of occupants, nest structure, and stage of development and to determine how they should be processed. Some are dissected immediately and identified on the basis of nest characters and scatology. Many are held for emergence to obtain species identifications of bees, their associates and nest competitors. Sticks are dissected after their occupants have emerged, and remaining contents are analyzed to determine survival rates and mortality factors that can be assessed within the limits of our methods.
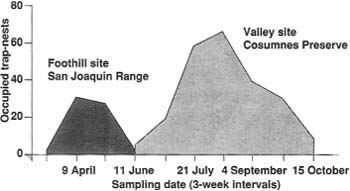
Fig. 2. Seasonal pattern of nest occupation by cavity-nesting bees in block sampling units (BSU) at San Joaquin Experimental Range and Consumnes River Preserve in 1990.
Swarm traps constructed of wood pulp and baited with Nasonov pheromone are monitored for establishment of honey bee colonies to determine reproduction of feral honey bees. With examination of the composition of fecal pollen, established colonies are subjected to detailed analyses of adult populations, brood, stores of pollen and honey, amount of comb, brood cell size and floral resource use. Some are monitored for survival for 1 or more years.
Preliminary results
Diversity, phenology, and relative abundance of both flowering plant and bee populations are similar among the three valley sites; foothill sites compared with valley sites differ. The BSU data show that in foothill sites most bee nesting activity occurs from early March to late May; in valley sites most nesting occurs from late May to mid-October (fig. 2). Megachile rotundata Fabricius is apparently most abundant in early summer, while M. apicalis Spinola increases through summer into autumn. Nesting of solitary bees at CR has declined since 1987.
Occupancy of nest holes. Most of our block-sampling units (BSU) were occupied by leafcutting, mason, resin and cotton bees (family Megachilidae, see table 1). The native orchard mason bee, Osmia lignaria propinqua Cresson, dominates BSU at the foothill sites. Introduced leafcutting bees, Megachile (Eutricharaea), dominate the valley sites, but there are habitats within these sites where native megachilids are abundant. Other arthropods occupy the BSU including cavity-nesting wasps, earwigs, ants, weevils, moths and spiders. European earwigs occur locally or seasonally in large enough numbers to reduce bee nesting.
Preference for hole size. Although larger holes have been offered, bees at the study sites nest primarily in 4.7- to 7.8-mm diameter holes. Selection of hole size varies. Females of M. rotundata, for example, tend to occupy the smallest diameter holes, while females of M. apicalis are more flexible. In late summer and autumn, at peak population, M. apicalis will accept larger diameter holes, constructing cells in a spiral or side by side in tunnels over 9 mm in diameter. This species will also accept holes in the ground, such as those from which Diadasia enavata (Cresson) emerge.
Mortality sources. Mortality factors, as determined by our rearing records, X-rays and nest dissections, include chalk brood fungus, parasitoid wasps, cuckoo bees and nest-destroying beetles. Earwigs, ants and other cavity users may also disrupt or damage bee nests. Supersedure, or nest take-over, has been found between females of the same species and between diverse species of cavity nesters. This may result in mortality from destruction of the previous brood by nest-usurping females and/or from emergence by earlier developing individuals through cells containing younger broods. Supersedure is, overall, relatively infrequent, but it may be locally or seasonally frequent.
Honey bee reproduction. The continuous high rate of swarm captures at CO contrasts with decreased swarm captures at CR from 1988 through 1990 and lack of swarm captures at SJ. At the valley sites, swarming occurs mostly in April. Occupation of swarm traps and survival of newly established colonies are best at riparian sites. Colonies may survive more than 1 year inside swarm traps, but those constructing nests externally on the swarm traps tend not to survive winter.
Feral EHB. Observations of feral EHB colonies at SL and CO demonstrate that they can survive for more than 4 years in suitable habitats. Small colonies, especially in ground cavities where size is constrained, are vulnerable to predation by small mammals or ants and may not survive the first year.
Discussion
Each study site is large enough to contain several habitat types. The great similarities noted in bee and flower communities among the three valley sites are not surprising, since they are usually classified in the same plant climate, plant community and faunal community types. The foothill sites also resemble each other in major plant and faunal community types, but they contain more distinct biotic elements, vary more in topography and differ most from valley sites. Early bee activity at the foothill sites is related to the flush of bloom in early spring, followed by dry summers. In contrast, the summer-autumn peak of activity in the valley sites corresponds to the peak bloom of Asteraceae.
It is difficult to monitor bee populations due to mobility of foraging bees, differences in degree of host specificity to pollen resources and difficulties in locating nest sites. Activities and relative abundance of selected bees are monitored regularly, using BSU and swarm traps for cavity-nesting bees and honey bees, respectively. Feral honey bee colonies and ground-nesting solitary bees are also monitored as nests are encountered. Measures of visitation to selected flowers are used to assess these and other bees whose nests may not have been located. Using these methods allows us to determine whether there is potential competition for food, nest sites, both or neither. The 3-week intervals at which BSUs are collected were designed to prevent saturation, but were considered long enough to obtain adequate frequency counts of bee nests to make quantitative comparisons of abundances between years and sites.
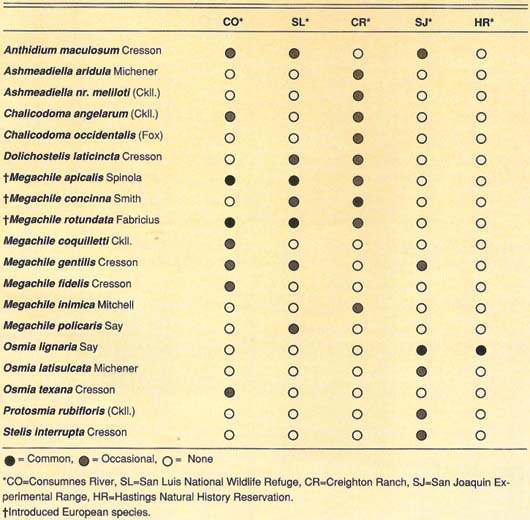
Some believe that M. apicalis may displace its close relative, M. rotundata, through competition for nest holes. Megachile rotundata appeared in California by 1949 and M. apicalis by 1982. Our data show that there is no complete overlap in seasonal abundance or of hole sizes used for nests. This suggests that these bees may coexist by partitioning the habitat with M. rotundata nesting early and in small holes, while M. apicalis dominates in late summer and autumn and is more flexible in selecting hole sizes. Both species are abundant in our BSU monitoring program, thus providing us with an opportunity to study invasion ecology of these solitary bees and their effects on native bees and wildland environments. Our observations on flower visitation and nesting behavior suggest that M. apicalis could be managed for pollination of commercial sunflowers, as is M. rotundata on alfalfa seed crops.
Because the BSU are only in the field for 3 weeks, they are not exposed to all potential sources of mortality. Many parasi-toid wasps, for example, attack bees only after they have spun their cocoons; few bees have developed to this stage by the end of 3 weeks. Thus, the standardized sampling interval adopted in 1990 is not sufficient to monitor accurately this type of mortality.
Five years of drought seem to have severely depressed the abundance of cavity-nesting bees, including honey bees, especially in the most southerly and least riparian sites (CR and SJ). To assess the magnitude of drought effects will require long-term monitoring of these populations, as several years of increased rainfall may be needed for them to respond; feral honey bees, however, may rebound more quickly.
Conservation biologists are aware that the ecological impacts of introduced species are well documented for birds and mammals, but that documentation for bees is inadequate. This lack is surprising in light of the importance of bees as pollinators for the reproduction of plants and for the production of food for vertebrate wildlife. Evidence is building that native bees are needed to maintain native flora, including rare and endangered plants, and that sound conservation policies for wild-land areas will be needed to include protection of bees and their nesting habitats. Our long-term studies will provide basic data for policy decisions about conserving flora and bees.
We have identified some basic components of the bee community structure, emphasizing cavity-nesting bee species, their competitors and floral resources. Community level studies of bees are lacking. The revitalization of community ecology through methods of ordination analysis allows interpretation of complex data sets with several variables, and provides us with new ways to learn which factors determine bee community structure. With this framework, we are now able to pursue studies that will accurately measure the ecological consequences of the arrival of AHB in California.
Ecological research: Long-term studies to gauge effects of invading bees

