


All Issues
Proximity to citrus influences Pierce's disease in Temecula Valley vineyards
Publication Information
California Agriculture 55(4):13-18. https://doi.org/10.3733/ca.v055n04p13
Published July 01, 2001
PDF | Citation | Permissions
Abstract
Pierce's disease has caused extensive losses to grapes in the Temecula Valley. The primary vector of Pierce's disease in the region is the glassy-winged sharpshooter (GWSS), which has been found in large numbers in citrus trees. We examined the role of citrus in the Temecula Valley Pierce's disease epidemic and found that citrus groves have influenced the incidence and severity of Pierce's disease in grapes. Because GWSS inhabit citrus in large numbers, California grape growers should take additional care with vineyards located near citrus groves.
Full text

Pierce's disease has caused significant losses to the grape industry in Temecula Valley. The region's mix of grapevines and citrus, which can harbor the glassy-winged sharpshooter, may be a factor in the disease's spread.
The grape industry in Temecula Valley of Southern California has experienced significant losses to Pierce's disease since it was first documented in 1997. This disease is caused by a bacterium Xylella fastidiosa Wells, which reproduces in the xylem and plugs the water-conducting tissue of the grapevine. Infection causes a chronic and progressive water stress condition that eventually kills the plant. The bacterium is vectored by sharpshooter (Cicadellidae), several species of which are found in the Temecula Valley.
The increase in incidence of Pierce's disease is chronologically linked with observed increases in numbers of the glassy-winged sharpshooters (GWSS), Homalodisca coagulata (Say) (Blua et al. 1999; Purcell and Saunders 1999a).
This sharpshooter is an invasive species in California; it was first detected in the state in 1989 (Sorensen and Gill 1996). The GWSS is native to the southern United States and northern Mexico and probably was introduced into California on plants from those regions.
Presently, there is a fundamental lack of understanding about the epidemiology of GWSS-transmitted Pierce's disease in the Temecula Valley. The epidemiology of plant diseases caused by arthropod-vectored pathogens is dependent on the abundance of vectors and pathogen sources, and the proximity of these to the plant host. For example, in San Joaquin Valley, Pierce's disease is sparsely distributed and spatially associated with pastures or weedy alfalfa, both of which include grasses that harbor important disease vectors in that area (Hewitt et al. 1946; Purcell and Frazier 1985).
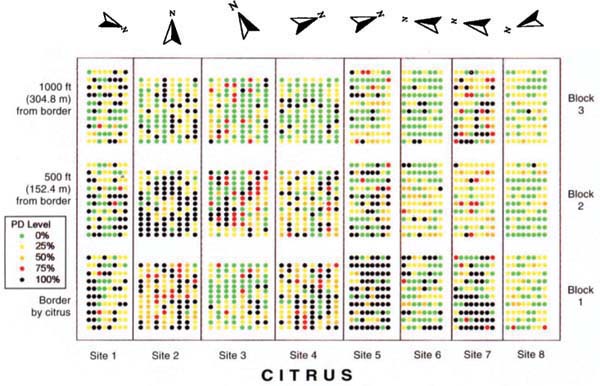
Fig. 1. Pierce's disease incidence from three blocks (0, 500 and 1,000 feet from citrus) in eight Temecula Valley sites, September 2000. Each dot represents the rating from one quadrilateral of a single vine.

TABLE 1. Average symptom severity among grapevines in eight Temecula Valley vineyard sites at three distances from citrus, September 2000

TABLE 2. Percentage of symptomatic plants in eight Temecula Valley sites within blocks at three distances from citrus, September 2000
Piérce's disease in California's north coastal region is most severe close to riparian areas where the primary vector, the blue-green sharpshooter, overwinters (Purcell 1974, 1975). Vineyards that are located 400 to 600 feet from riparian areas typically do not experience damaging levels of Pierce's disease (Purcell 1974).
Since the introduction of GWSS in Temecula Valley, the epidemic has spread as rapidly and extensively as it did in the southeastern United States, where GWSS-transmitted Pierce's disease is the major factor preventing grape production (Purcell 1981). An epidemiological study conducted in that area showed that phony peach disease, which is caused by another strain of GWSS-transmitted X. fastidiosa, diminished with distance from local sources of disease. However, the effect of local sources extends to nearly 1,000 feet, instead of the 400 to 600 feet of vineyard that have been affected by local sources of Pierce's disease in California (KenKnight 1961).
Other work has shown that GWSS dispersal is only loosely related to its local reproductive source (Ball 1979). California grape-growing regions already produce many different plant resources that can be used by GWSS as feeding and reproductive hosts (Adlerz 1980; Blua et al. 1999) and by X. fastidiosa (Freitag 1951; Hewitt et al. 1946; Hopkins and Adlerz 1988; Purcell and Saunders 1999b).
The key question for the California grape industry is whether it is possible to manage GWSS-driven Pierce's disease epidemics, which are likely to be less localized than anything experienced here in the past. Our understanding of the spatial relationships among vineyards, other sharpshooter's reproductive hosts and sources of Pierce's disease, probably no longer applies now that GWSS has been introduced. Until these new relationships have been described under local conditions, there is no reason to believe that a grower will be able to prevent Pierce's disease infection by initiating mitigating measures within a single vineyard. We attempted to determine the current role of citrus — an excellent GWSS host — on Pierce's disease incidence and severity in California.
Vineyard survey
We selected eight vineyards for our survey that met all of the following criteria:
-
At least one side bordering citrus.
-
Continuous grapes, a minimum of 1,000 feet from the citrus.
-
Large enough that blocks 500 feet and 1,000 feet from the citrus were at least that far, respectively, from any other border of the vineyard, in order to determine whether the disease gradient was related solely to citrus. At the furthest distance surveyed, we wanted the grapes to be surrounded only by other grapes.
-
A single grape variety within each vineyard site, in order to compare the incidence of Pierce's disease within the vineyard by visual symptoms (symptom severity varies with grape variety).

The invasive, nonnative, glassy-winged sharpshooter transmits the bacterium Xyllela fastidiosa, which causes Pierce's disease. Young vines in Temecula Valley show symptoms at the base of several canes (stems).
Within each of the eight vineyards, we selected three blocks for sampling, each block measuring as close to 100 feet by 100 feet as was practical, considering local row and vine spacing. Block 1 was at the border of the vineyard adjacent to citrus. The middle block (2) was centered at 500 feet from the citrus border, and the distant block (3) extended up to 1,000 feet from the citrus border along a transect perpendicular to the border (fig. 1).

The study found that proximity to citrus influenced the incidence and severity of Pierce's disease in grapes.
Pierce's disease symptoms were rated for every vine in each block on a scale of 0,1, 2,3 or 4, representing healthy, 25%, 50%, 75% and 100% of the vine showing Pierce's disease, respectively. The differences in ratings were not qualitative, but quantitative, that is, the proportion of the plant showing symptoms. Symptoms of Pierce's disease in the late summer in Temecula include leaf marginal necrosis, early drying of the fruit (“raisining”), and “green islands,” which are areas of abnormally green bark surrounded by mature brown bark on the canes.

X. fastidiosa reproduces and plugs water-conducting tissues, causing water stress and killing the plant. In late summer and fall, grape foliage (white variety) infected with Pierce's disease becomes slightly yellow at the margins and then dies, leaving concentric zones of progressive discoloration. Photos by Jack Kelly Clark.
Prior to the study, we trained four samplers to recognize Pierce's disease symptoms; each sampler evaluated the same number of vines in each block. In order to confirm that disease symptoms were due to infection with Pierce's disease, representative samples were tested for the presence of X. fastidiosa using a pathogen-specific diagnostic technique, ELISA (enzyme-linked immunosorbent assay) This technique detected X. fastidiosa in 66%, 81% and 86% of the samples rated 1,2 end 3 respectively.
There are several reasons why ELISA may not detect X. fastidiosa in infected plants, including assaying segments of infected plants that did not contain bacterial cells and its limitation in detecting small quantities of X. fastidiosa in infected plants. These explanations are supported by the fact that detection by ELISA increased with symptom severity. As described by Varela et al. (2001), symptom expression provides a reliable indicator of infection.
We conducted a complete Pierce's disease census in each block. In order to satisfy the assumptions of an analysis of ANOVA, we randomly selected 60 plants per block for analysis. Disease ratings were arcsine-transformed prior to analysis (Sokal and Rohlf 1998). With each site representing a replicate, we used an analysis of variance to detect differences in Pierce's disease levels as a function of distance from the edge of the vineyard adjacent to citrus. After we analyzed the ratings, we converted them to percent infection for display purposes. We also analyzed severity data to determine the spatial relationship of disease within each of the blocks, with respect to the distance from citrus. For this analysis, the disease ratings were arcsine-transformed and then regressed on each vine's distance from the border of the block closest to the citrus (SAS Institute 1999).
For a second series of analyses, we classified all infected plants in one class and healthy plants in another class. These data were subjected to a chi-square test for independence, which indicated whether there were statistical differences between the blocks in each field (Sokal and Rohlf 1998). A final test evaluated the relationship between the number of infected plants in each block and the block's distance from the citrus.
A matter of proximity
The average level of the expression of Pierce's disease symptoms for vines evaluated in the eight fields varied greatly, from 9.4% in site 8 to 61.7% in site 2, with an average across all sites of 37.2% (table 1). Average disease ratings for vines in the 24 blocks varied even more, ranging from 5.1% to 90.2%. In four sites (1, 4, 5 and 6), there was an apparent gradient from highest average percent infection near citrus to lowest average percent infection far from citrus (table 1; fig. 1). In two of the sites (2 and 3) the highest average infection was in the center block, while two sites (7 and 8) had relatively the same average infection in blocks closest and furthest from citrus, with less infection in the center block. Analysis of variance of disease ratings from all eight vineyards showed statistically significant (P < 0.05) differences in disease severity between the three distances from citrus. This analysis indicates that Pierce's disease infection in Temecula Valley is most severe when the vines are adjacent to citrus, and that the damage declines as one moves away from citrus. At none of the sites was the severity of Pierce's disease inversely related to proximity to citrus.
On a smaller scale, an analysis of disease gradients within each of the 24 blocks showed five blocks (site 2, block 2; site 3, block 1; site 6, block 2; site 7, block 1; and site 8, block 3) with statistically significant (P < 0.05) relationships showing higher disease severity closer to citrus. Two of the blocks (site 1, block 3; and site 8, block 2) had significantly (P < 0.05) higher disease furthest from citrus. Gradients within these blocks are easily visualized (fig. 1). The other 17 blocks showed no significant gradient with respect to distance from citrus. For all blocks the R-square values were quite low, ranging from near zero to 0.12. The R-square, or the coefficient of determination, measures the proportion of variation in the data that can be explained by some variable, in this case distance from citrus. The low R-square indicates that no more than 12% of the variation in distribution of Pierce's disease within individual blocks can be attributed to distance from citrus. These analyses highlight the limitation of using small blocks to evaluate the spatial distribution of Pierce's disease spread by GWSS.

In a red grape variety infected with Pierce's disease, leaf margins become slightly red and die. Photos by Jack Kelly Clark.
The number of plants showing symptoms (all levels of severity) in each vineyard ranged from 51.1% to 87.1% (table 2). Sites 3,6 and 8 had 50% to 60% diseased vines while sites 1,2,4,5 and 7 had percentages of diseased plants between 70% and 88%. When the data within each site were analyzed, the gradient of Pierce's disease did not consistently decrease with distance from citrus. At three of the eight sites (sites 2, 4 and 5; table 2), the number of diseased vines exhibited a significant linear trend away from citrus (P < 0.005), indicating that citrus was important to Pierce's disease incidence in these fields. A similar trend (although not statistically significant) was shown at site 1. At sites 3 and 6, the largest percentage of diseased plants were in the middle block (P < 0.003). At site 7, the three blocks had similar numbers of diseased plants, and at site 8, the blocks closest and furthest from citrus had the highest levels of disease (P < 0.01), with a lower number of diseased plants in the center block (table 2).
Epidemiological applications
This study yields valuable information about the GWSS-vectored Pierce's disease epidemic in Temecula Valley, the first such epidemic in California. First, we described the spatial distribution of Pierce's disease at two different scales. Such analyses are necessary to understand the dynamics of GWSS-transmitted Pierce's disease epidemics. Our work at the larger scale suggests that the effects of local GWSS-sources on X.fastidiosa spread should be studied with transects of at least 1,000 feet; even at this distance we found high levels of disease. This is considerably greater than the gradients determined for Pierce's disease transmitted by the blue-green sharpshooter near riparian areas of Northern California (Purcell 1974). At the smaller scale, we discovered the lack of a discernable gradient of Pierce's disease within the 100-foot-by-100-foot blocks, suggesting that studies of GWSS movement and Pierce's disease spread will not be productive at small scales. Clearly, growers and researchers should consider spatial scale when designing management strategies to reduce the spread of Pierce's disease. They also should consider scale when establishing sampling plans to answer questions about GWSS distribution and disease spread.
Second, our data indicate that proximity to citrus has influenced the incidence and severity of Pierce's disease in Temecula Valley. When we used sites as replicates and analyzed for differences in severity between blocks, we found significantly higher disease severity closest to the citrus, with lower severity further from citrus. Additionally, when the vines were classified as healthy or infected, we found statistically significant linear trends away from citrus in three of the eight sample sites.
Much of the vineyard acreage in Temecula Valley lies within 1,000 feet (38%) and 2,000 feet (72%), respectively, of citrus groves (determined by a buffering operation applied to grape and citrus locations [ESRI 1999]). It is likely that this has been a factor contributing to the rapid dissemination of X. fastidiosa throughout the valley. However, just as proximity to citrus was related to disease gradients in some vineyards, disease in other vineyards could not be explained by proximity to citrus; other variables were involved. To establish a reliable estimate of the spread of the disease, we must begin collecting data early in an epidemic, when the number of diseased plants is limited, and continue as the infection spreads.
By the time we surveyed in Temecula Valley, so many of the plants were infected that gradients from adjacent citrus might have been masked by the spread of Pierce's disease related to other factors, such as proximity to other GWSS or Pierce's disease hosts, soil condition, irrigation or fertilizer. For example, two vineyards (sites 2 and 3) had higher disease incidences in the middle blocks (table 2), suggesting that variables other than citrus proximity influenced the expression of Pierce's disease symptoms.

As the infection spreads, the top of the vine dies while scion or rootstock suckers sprout from near the soil level. Grape growers throughout California should be especially watchful of vineyards located near citrus groves, or near other possible, noncitrus glassy-winged sharpshooter hosts.
Alternatively, these findings may also contribute evidence that vines in the center blocks became infected prior to vines in the other two blocks, and X. fastidiosa was spread from these early infection sites to surrounding plants. Disease severity data, which provide an indication of the duration of infection (assuming the longer a plant is infected, the more severe the disease) showed greater severity in these two middle blocks. Such secondary spread of Pierce's disease, from vines within the vineyard to neighboring vines, has not been documented in California, but may occur when GWSS is the vector. If so, chemicals that interfere with acquisition of X. fastidiosa from infected vines or transmission to healthy vines may reduce the spread of the pathogen, the way chemicals interfere with persistently transmitted arthropod-vectored viruses (Perring et al. 1999).
While the association between citrus and the spread of Pierce's disease in grapes will need to be examined statewide, this study suggests that grape growers throughout the state should be particularly watchful of vineyards near citrus groves. Similarly, other areas containing noncitrus GWSS host plants (see http://plant.cdfa.ca.gov/gwss/ for GWSS hosts) should be monitored for sharpshooters, and vineyard borders near these areas should be surveyed for Pierce's disease.
Combining insecticides for shortterm control with other long-term strategies such as removing local sources of GWSS, maintaining spatial distances between vineyards and sources, and planting new vineyards far from vector sources may also help to reduce the spread of X. fastidiosa vectored by GWSS.
Proximity to citrus influences Pierce's disease in Temecula Valley vineyards

