


All Issues
Special Section Insert: Lessons from a record-breaking freeze: Some olives show damage; many, coldhardiness
Publication Information
California Agriculture 47(1):1-12.
Published January 01, 1993
PDF | Citation | Permissions
Abstract
A once-in-a-century cold front, expressed as an advective freeze, damaged ‘Manzanillo’ extensively statewide. ‘Ascolano,’ ‘Sevillano’ and ‘Mission’ received minor damage. Damage included tip burn, defoliation, bark splitting and limb dieback. The next growing season some new leaves were deformed and flower bud damage could be found. Outbreaks of olive knot had also been expected, but few occurred.
Full text


‘Manzanillo’ trees in an orchard near Visalia show the extensive freeze damage characteristic of young trees.
During the last 2 weeks of 1990, a mass of dense, frigid Arctic air moved into California, and temperatures plunged to record lows along the Pacific Coast and in the Central Valley. Not only were temperatures record breaking, but the cold spell's duration and its widespread effect were noteworthy. This “Arctic Express,” as it was called by the media, was a once-in-a-lifetime experience for most Californians accustomed to mild winters. Once immediate concerns were dealt with, such as frozen water pipes, attention turned to the cold front's effects on California's agriculture. One major crop adversely affected was olive (Olea europaea L.). Responding to concerns of olive growers, the California Olive Committee (COC) requested the University of California's Department of Pomology at Davis to study the olive under freeze conditions.
EDITOR'S NOTE:
Late in December 1990, a once-in-a-century cold front moved through California, threatening olive, citrus and nursery plantings across the state. Responding to olive growers’ concerns, the California Olive Committee funded a UC study of olive freeze damage – yielding a wealth of data on olive performance under exceptional freeze conditions. Printing of this special section was funded by the California Olive Committee.
Cover photos illustrate olive damage seen throughout the state after the December 1990 freeze.
The study has been broad based, from botanical and horticultural aspects of freeze damage and recovery to climatic and economic questions. Our goals were: (1) to understand the nature of freeze damage in olive, (2) to explore the question of olive cultivar coldhardiness, (3) to learn the frequency of such climatic events in California, (4) to estimate the potential economic impact on producers and the state economy, and (5) to develop for California's olive producers guidelines for minimizing the effects of future freezes.
Our study included (1) field examination of olive orchards, (2) microscopic examination of freezed-damaged olive tissue, (3) laboratory experiments to look at coldhardiness in California cultivars, (4) climatological research, (5) questionnaires sent to olive growers and (6) economic studies.
Field work was carried out in California's two major olive-producing regions: Butte, Tehama, and Glenn counties in the Sacramento Valley and Tulare and Madera counties in the San Joaquin Valley. Assisting were UC Cooperative Extension farm advisers and olive growers. The COC mailed approximately 1,400 questionnaires regarding the freeze to growers; 311 responses were received and used for data processing and analysis. The questionnaire dealt not only with damage symptoms and severity but also with production levels and cultural practices.
December 1990 and olive climates
Early in the third week of December 1990, a low-pressure trough moved through California bringing light precipitation. Behind the trough came cooler air that caused temperatures to drop, followed shortly by a large mass of dense, frigid Arctic air. By early morning on December 18, most of the Central Valley experienced freezing temperatures. The air mass continued to build and, on the nights of December 22, 23 and 24, temperatures plummeted. New record minimum temperatures were established. Subsequent damage to olive occurred with this combination of cold and its duration: Central Valley cities stretching from Redding to Bakersfield recorded lows below 20°F (-6.7°C) on these first 3 days of winter, and freezing nighttime temperatures persisted into the first week of the New Year.
Record low temperature data for Davis and for several olive-growing sites in the Sacramento and San Joaquin valleys are given in table 1. The December 1990 freeze set new record lows for all sites, except Davis and Visalia. At some sites temperatures were possibly either lower or higher than those given in table 1. For example, official National Climatic Data Center (NCDC) records show Visalia's lowest temperature was 21°F (-6.1°C) on December 24, but data gathered by the California Irrigation Management Information System's (CIMIS) automated station in Visalia show the lowest temperature was 17.3°F (-8.1°C) on December 23.
Low temperatures were accompanied by persistent winds, bringing about a condition known as an advective freeze. Because wind mixes air so that colder, dense air has no chance to accumulate at ground level or to drain to low-lying areas, these winds may have prevented even lower temperatures. Advective freezes also have a very low dew point (the temperature at which air becomes saturated and produces dew). The lower the dew point, the more potentially damaging. The lowest dew points of interest recorded by CIMIS during the 1990 freeze were 15°F (-9.4°C) at Visalia December 23, 0°F (-17.8°C) at Davis December 21, and -9°F (-22.8°C) at Orland, also on December 21.
Advective freezes may be contrasted with radiation frosts, which take place under still or very low wind conditions. Colder air accumulates near the ground; warmer air can be found at altitudes of 25 to 50 feet (7.6 to 15.2 m). Cold air is heavy and therefore migrates to valleys and other low-lying areas. In such frosts dew point is at freezing or a few degrees below it. Radiation frosts usually occur in early to mid-autumn or late winter to early spring. These frosts rarely seriously affect olive trees, although unharvested fruit may be damaged in autumn, as sometimes occurs in ‘Mission’ olives. Such frosts may damage other fruit crops, especially those characterized by early spring flowering.
Although temperatures at which olive trees can be damaged vary, depending on climatic conditions, temperatures at or below 20°F (-6.7°C) are often critical. The frequency of such episodes in California's olive-growing areas varies considerably (table 2). Generally, the San Joaquin Valley is less threatened by such freezes than is the Sacramento Valley. For example, Willows in Glenn County may see such temperatures once every 5 years, whereas Visalia in Tulare County may experience such freezes once every 17 years. Timing of such freezes also varies. The Sacramento Valley (Orland, Willows, Oroville, and Davis) is more susceptible to damaging freezes in early and mid-December before trees have had time to harden off than is the San Joaquin Valley (Visalia and Porterville) (table 3). Normally, cool, wet weather in November and December in the Sacramento Valley acclimates olive trees so that they can resist freeze damage – but not always. Such acclimation is normally less likely in the San Joaquin Valley. However, according to CIMIS data, by the time of 1990 freeze, Visalia in the south had received 198 hours ≤ 33.8°F (1°C), whereas Orland in the north had received only 111 hours of such temperatures.
Although new records were set at many California sites during the 1990 freeze, the Central Valley had previously experienced cold fronts. According to NCDC records, serious freezes in which temperatures dipped below 20°F (-6.7°C) occurred at several sites in 1913, 1919, 1930, 1932, 1937, 1948, 1949, 1950, 1972, and 1978. Of the freezes before 1990, the 1913 and 1932 freezes were the most severe and extensive (table 1). The frequency and geographical extent of such freezes varied. Temperature records before 1913 are sketchy for many sites in California, but what records exist indicate another extensive freeze, capable of damaging olive trees, occurred in 1888.
California olive-producing regions are not alone in experiencing damaging freezes. Olive trees in northern Italy have been damaged at least five times in this century, the most recently in 1989. Records for Italy also indicate at least three freezes in the 19th century and four in the 18th century. The olive-growing regions of Azerbaijan along the Caspian Sea also report several episodes of damaging temperatures. Trees were reportedly exposed to freezes of 17.6°F (-8°C) in 60% of the years studied, of 14°F (-10°C) in 40%, of 8.6°F (-13°C) in 5%, and of 3.2°F (-16°C) in 1%. Indeed, most northerly growing areas, from Spain in the west to China's Turkic regions in the east, are subjected fairly regularly to low temperatures that may damage olive trees. Freeze damage to olive is not always limited to the north. Defoliation and limb dieback have been observed on olive trees on the warm island of Crete as a result of a freeze during the winter of 1991–1992
Factors affecting coldhardiness
Cultivar differences
Although olive, as a species, is the most cold-hardy of the subtropical fruit trees, certain olive cultivars are more cold-hardy than others. For example, in the 1990 California freeze, ‘Manzanillo’ trees suffered more damage than did any other cultivars grown in the state. Where ‘Manzanillo’ was grown alongside ‘Mission,’ ‘Ascolano,’ or ‘Barouni’ in the same orchard, the ‘Manzanillo’ trees sustained obvious damage, but the other cultivars sustained little or none. Responses of growers to the questionnaire clearly support differences in coldhardiness among the five major California cultivars (table 4). ‘Manzanillo’ was the most damaged. Other cultivars appeared to be hardier; ‘Ascolano’ emerged as the hardiest. Table 5 provides an evaluation of the coldhardiness of olive cultivars from various countries held in the USDA collection at Winters and observed following December 1990. Italian scientists suggest that differences in olive coldhardiness may be related to the stomatal density on leaves of different cultivars with hardy cultivars having fewer stomata per unit of area.
Acclimation
Hardiness increases when trees are exposed to cold temperatures as autumn proceeds into winter, a process called “acclimation.” The importance of acclimation can be illustrated by an example of a mature ‘Manzanillo’ orchard in Glenn County, chosen for a study specifically because it had been damaged in a freeze in November 1985. As a consequence, defoliation, bark split, and olive knot infestation were so great that the trees were cut to within 2 feet (0.6 m) of the ground, and new scaffolds and canopies were developed from shoots that sprouted on each stump. Although temperatures reached only 22°F (-5.6°C) at nearby Willows during this episode, the experience came so early in the acclimation process that little hardening off had occurred.
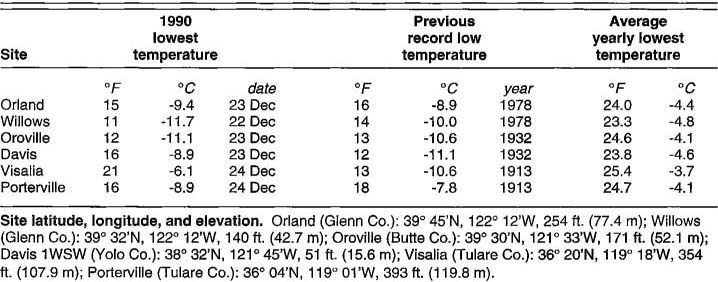
TABLE 1. Low temperatures recorded in California olive-producing regions,* from National Climatic Data Center, 1913–1990

TABLE 2. Number and frequency (%) of years reporting at least one occurrence of minimum low temperatures below given temperature thresholds in olive-producing regions in California, 1913–1990

TABLE 3. Timing of lowest yearly minimum temperature of ≤ 20°F (-6.7°C) in California olive-producing regions, 1913–1990
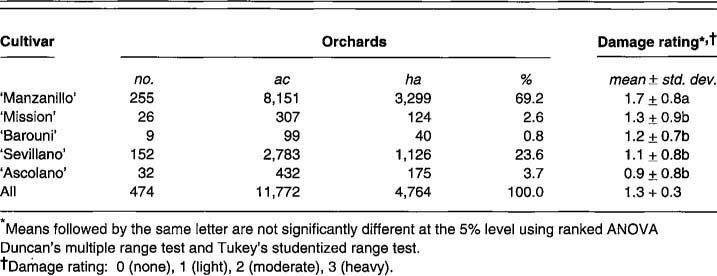
TABLE 4. Statewide freeze damage estimation for five olive cultivars as reported on grower questionnaires
![Subjective rating* of December 1990 cold (lowest temperature: 15°F [-9.4°C]) damage to olive varieties in the U.S. Department of Agriculture Germplasm Repository in Winters, California](http://calag.ucanr.edu/archive/?image=tab4701ps4b.jpg)
Table 5. Subjective rating* of December 1990 cold (lowest temperature: 15°F [-9.4°C]) damage to olive varieties in the U.S. Department of Agriculture Germplasm Repository in Winters, California

By contrast, in December 1990, the trees, now some 12 feet (3.7 m) tall, had had more time to harden off, and damage was relatively minor, even though the temperature reached 11°F (-11.7°C) at Willows. Increased tree age perhaps also contributed to the better outcome in 1990. Differences in damage to California cultivars from place to place may be partly explained by differences in acclimation (table 6). However, differences in cold severity also affect levels of damage. Acclimation differences cannot be used to explain the perceived greater extent of damage to trees in the San Joaquin Valley as compared with those in the Sacramento Valley because, as mentioned, Visalia in the south had, in fact, received more hours at acclimating temperatures than had Orland in the north. The greater total damage in the south is probably due to the extensive ‘Manzanillo’ acreage.
Cultural practices
Pruning, irrigation and fertilization affect tree growth and thus influence coldhardiness. For example, in a ‘Sevillano’ orchard near Corning, trees pruned in the fall before the freeze produced vegetative growth that did not have enough time to harden off before December's severe cold. Although ‘Sevillano’ trees are normally rather cold-hardy, pruned trees were damaged by the low temperatures; nearby unpruned ‘Sevillano’ trees escaped injury (fig. 1). This pattern was repeated in numerous locations statewide and with other cultivars. The open canopies on pruned trees probably exposed tree parts to frigid air and wind stress. Wind stress may also account for increased damage seen in trees along orchard perimeters.
Freeze damage was evaluated on ‘Manzanillo’ trees in a Madera County orchard in which UC researchers conducted irrigation trials. As measured by the extent of defoliation, damage was greater on trees receiving more water than on those receiving less. Excessive irrigation, especially when continued after harvest, may elicit more vegetative growth and may make trees more subject to damage. Conversely, limited water may promote tree desiccation, thereby increasing hardening off as well as retarding vegetative growth. In terms of tree recovery, however, well watered trees responded quickly and began strong growth once temperatures warmed in spring 1991; recovery of low-irrigation trees was less vigorous. Moreover, leaf loss was greater on high-irrigation trees immediately following the freeze, but defoliation from stress under high temperatures in summer 1991 was greater than on low-irrigation trees. Defoliation on all ‘Manzanillo’ trees was more extensive than normal throughout the growing season, but low-irrigation trees lost many more leaves as the season progressed.
The bottom line with respect to irrigation seems to be this: (1) little or no irrigation after harvest encourages hardening off before cold weather and decreases chances of freeze damage, and (2) adequate irrigation in spring encourages recovery from freeze damage.
Clear evidence of damage from fertilization did not emerge in our study. Nitrogen applied late in the season (after July), or in amounts or formulations that persist into fall, may account for continued vegetative growth that will not harden off before winter arrives. Trees, therefore, are more susceptible to damage from low temperatures.
Previous studies
What affects olive coldhardiness in California mirrors what has been noted in Italy. There, duration of freezing temperatures and tree age were listed as important factors - the same factors probably operative in California's 1990 freeze. Repeated freezing and thawing over more than a week no doubt contributed to physical stress and strain on trees. Growers’ responses to the questionnaire indicated that young trees were more severely affected than were older trees, regardless of cultivar (table 7).
Temperatures ranging from 19.4°F (-7°C) to -4°F (-20°C) have reportedly damaged olive. The 1932 freeze damaged Sacramento Valley olive trees at 17°F (-8.3°C); others were not damaged until temperatures dropped to 11°F (-11.7°C). In the former Soviet Union, temperatures from 17.6° to 14°F (-8° to -10°C) slightly damaged olive trees, whereas temperatures of 1.4° to -7.6°F (-17° to -22°C) killed trees down to the ground. Damaging temperatures are sometimes given as known thresholds.
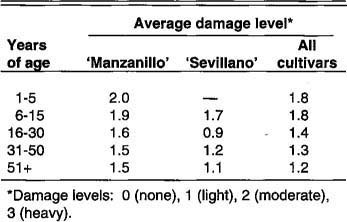

Fig. 1. This ‘Sevillano’ tree in Tehama County was pruned in November before the freeze and was severely damaged because of a loss of hardiness. The damage was made more severe by an infestation of olive knot disease.
The critical temperature parameters given here are based largely on field observations, not on experimentation. This partly explains the reason for the great range in temperatures suggested as damaging to olive. Controlled laboratory experiments on the freezing characteristics of olive are limited. In one study using cold-acclimated olive tissues, 50% injury was seen on leaves at 10.4°F (-12°C), on leaf buds at 8.6°F (-13°C), on stem cambium at -4°F (-20°C), and on stem xylem at -0.4°F (-18°C). Experiments in Japan on an unnamed cultivar of olive, as part of a mass screening of more than 100 species, revealed damage to leaves and buds at 8.6°F (-13°C) and damage to twigs at 5°F (-15°C).
Laboratory experiments
We examined coldhardiness in California olive cultivars in laboratory experiments at the Washington State University Tree Fruit Research and Extension Center in Wenatchee, Washington. Samples were processed within 30 hours of harvest. Sampling was repeated at 1-month intervals beginning October 28, 1991 and ending April 13, 1992. Leaves of these cultivars were processed beginning in December rather than in October. Samples of shoots from ‘Ascolano,’ ‘Sevillano,’ and ‘Barouni’ cultivars were analyzed, along with ‘Manzanillo’ and ‘Mission,’ once on January 27, 1992.
In the Wenatchee laboratory, basal internodes and leaves of the current season's growth were cooled at a rate of 27°F per hour (15°C/hr) from room temperature to -40°F (-40°C) to determine the differential response of each sample. In this type of analysis, the target tissue's internal temperature is measured. Freezing events are noted by a rise in temperature as the latent heat of the fusion of water is released; this is called an “exotherm.” Computer-assisted collection of sensor signals was performed at a frequency of 20 Hertz (Hz).
The onset of freezing events under laboratory conditions may or may not indicate the survival temperature of a particular tissue under field conditions. Because of the great proportion of water in plant tissue, freezing always severely challenges the viability of affected cells. It cannot be assumed, however, that freezing will inevitably kill frozen tissue or the entire plant. To associate freezing temperatures with survivability, on two dates plant tissue samples were cold-treated to noted exotherm temperatures in separate experiments, then exposed to viability tests, and further observed over extended periods to assess plant survivability or functionality. In the viability tests on olive tissue, a modified form of the tetrazolium chloride (TTC) procedure described for apple was used.
Thermograms of the freezing of olive tissues showed from one to three exothermic peaks, depending on tissue morphology and the season. Each peak corresponded to the freezing of distinct cell aggregates. The area under each peak was proportional to the quantity of cell water and/or cell mass involved in the transition from water to ice.
In a typical olive shoot, the bark is composed of the cork cells of the exterior and the living phloem cells inside (which transport sugars and other substances to organs that need them). The wood is composed of xylem cells (which transport water and minerals from the roots), ray parenchyma cells (which relate the xylem to the phloem), and the pith (an area of undifferentiated cells at the center). The area between the xylem and the phloem is the cambium. The cambium is the source of all new cells, whether xylem or phloem, for lateral growth. In most species, freezing of pith cells almost always kills them, but does not impair plant survival. In contrast, freezing of xylem parenchyma or ray parenchyma cells kills these cells and the entire branch or shoot. Freezing kills any actively growing tissues. The first exotherm seen in olive stem tissue is that of the extracellular water and of bark cells. The second exotherm is that of the xylem ray parenchyma cells in the wood. A third exotherm has been observed only in stem tissue at the time of and after the onset of growth in spring. The third exotherm apparently has no connection with plant survivability, as indicated by the TTC viability test; however, it may be useful as a physiological marker of the beginning of growth. Leaf tissue exhibits one exotherm.
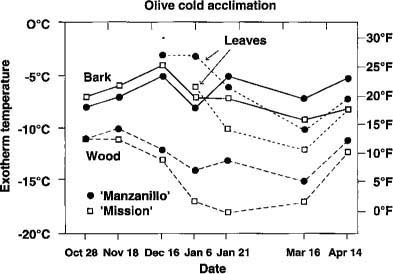
Fig. 2. The freezing temperatures for the bark as winter progressed do not indicate a capacity for acclimation, whereas the freezing temperatures for the wood and leaves show an unmistable pattern of acclimation and deacclimation for both of these cultivars.

Fig. 3. Tip burn (here on ‘Barounl’) is the death and blackening of tissues at or near the succulent growing tips. This breaks apical dominance, and two or more axillary buds will begin growing in the spring, thereby creating a bushier canopy.
In the current year's olive shoots, freezing in the bark accompanied freezing of extracellular water at temperatures well above such events inside the xylem and pith cells of the wood. Viability tests show that bark cells did not survive this freezing. Bark cells did not exhibit any acclimation as the season progressed (fig. 2). The fact that bark cells froze at higher temperatures than wood cells may be a significant factor by itself in determining the extent of damage to olive trees. Once it froze, the bark became rigid. The later freezing, in turn, of xylem water and xylem cells caused a volume expansion leading to splitting of the bark. This splitting could then lead to desiccation of bark and wood. The split could also be a wound allowing for entry of various pathogens.
Not only did the xylem and pith cells of the wood freeze at much lower temperatures than did the bark cells and extracellular water, but the temperatures of freezing in the xylem and pith cells also varied according to the time of season in a pattern, suggesting acclimation capability (fig. 2). This acclimation capability was also borne out by the fact that the xylem and pith cell nucleation temperatures could be significantly lowered by storage of unacclimated tissue in a moist environment at 36° to 40°F (2.2° to 4.4°C) for 7 days. Viability tests indicated that this freezing was associated with the death of these xylem tissues and that the consistently lower nucleation temperatures for ‘Mission’ xylem, compared with ‘Manzanillo’ xylem, mean that ‘Mission’ is a hardier cultivar. The one-time test on January 27, 1992 of ‘Ascolano,’ ‘Barouni,’ and ‘Sevillano’ with ‘Manzanillo’ and ‘Mission’ confirmed the existence of cultivar differences in coldhardiness (table 8). ‘Barouni’ and ‘Sevillano’ showed xylem freezing similar to that seen in ‘Mission.’
Remarkably, leaves showed a pattern of acclimation similar to that seen in the xylem (fig. 2). However, leaves are considerably less hardy than the xylem. At the beginning and end of the winter cold period, leaves were also less hardy than the bark. ‘Mission’ leaves were consistently hardier than ‘Manzanillo’ leaves. Especially in early season, leaf vulnerability to freezing at temperatures above that necessary for bark freezing may be an important factor in overall tree damage, as freezing events begun in the leaves may propagate into the stem. Leaf loss from freeze damage is also serious because leaves are important in the initiation of inflorescences and in the nourishment of developing fruits and other organs, and because leaf scars allow for olive knot infection.
Rootstock effects
The use of rootstocks can affect the coldhardiness of the scion graft to them in many fruit crops. Most olive cultivars in California are grown on their own roots, but numerous older trees in the Sacramento Valley have been grafted over to other cultivars, such as ‘Manzanillo’ or ‘Sevillano’ on ‘Mission.’ No information on the effects of rootstocks on coldhardiness emerged in field observations or from responses to the growers’ questionnaire. In the case of ‘Sevillano’ on ‘Mission’ rootstock, this is not surprising because these two cultivars are close to each other in degree of coldhardiness.

TABLE 8. California olive cultivar coldhardiness as indicated by bark and wood freezing temperatures in differential thermal analysis
Types of freeze damage observed
Some symptoms of damage to olive trees appeared soon after the December 1990 freeze. Other symptoms were expressed later. Damage symptoms by cultivar, as reported on the grower questionnaire, are summarized in table 9. Aspects of damage observed in the field included the following.
Tip burn
Tip burn refers to the death and blackening – as if by burning – of succulent, growing, shoot tips and nearby leaf tips. Tip burn was seen on all cultivars. On ‘Barouni,’ tip burn was the only readily apparent symptom of freeze damage in field observations (fig. 3). The growing tips consist of the meristematic region at the apex where cells are rapidly dividing to provide tree growth. The shoot differentiates and begins to mature in the region below the meristem. These areas are thoroughly hydrated and inherently unhardy. The abundance of free water in these tissues and their location on the perimeter of the tree canopy make them susceptible to freezing and the subsequent expansion that breaks surrounding tissue.
Tip burn often affected the first four or five nodes of the shoot. At the node below the area affected by tip burn, lateral buds broke, once growth started in spring 1991. Because the olive has opposite leaf and axillary bud placement, this resulted in the bifurcation of the shoot and an increased bushy appearance of the canopy as foliage matured. The increased number of shoots in the outer shell of the canopy led to an overproduction of inflorescences for the 1992 growing season. This could exacerbate the tendency towards alternate bearing found in certain California olive cultivars, especially ‘Manzanillo.’
Bark, wood, and leaf discoloration
The normal greenish grey appearance of young olive bark was darkened. The darkening may be the result of oxidation of phenolic compounds, spilled into cell walls when the membranes of living cells were disrupted by ice crystals. Normal olive wood is a cream color, but wood of damaged trees was stained in colors ranging from buff to dark brown. We were unable to use wood color to gauge the extent of damage to the whole tree because wood darkening was mainly localized in areas of damage.
Defoliation
Loss of leaves was the best overall indicator of the extent of damage to olive trees. These flat organs have much surface area exposed to the surrounding air, and they lose heat rapidly in response to dropping temperatures. Deciduous trees lose their leaves before winter, thereby avoiding damage from freezing. However, this strategy is not available to olive trees because they are evergreens. In many cases, bark split (discussed later) was seen directly below split leaf scars left after defoliation. As with the stem, leaves harden off as they are exposed to colder temperatures, as shown by our studies using differential thermal analysis (fig. 3). However, leaves are considerably less hardy than stems. Ice formation in leaves may not always lead to immediate defoliation, as is discussed in a section on leaf chlorosis.
Defoliation is part of the natural process known as “abscission” which is mediated by the gaseous plant hormone, ethylene. Normally, olive leaves persist on the tree for 2 or 3 years when an abscission zone forms in special cells on the petiole, or leaf stem, and the leaf drops away. Olive leaves may drop at any time of the year, but most do so just before the resumption of growth in spring.
Defoliation was seen to one extent or another on all cultivars as a result of the 1990 freeze. ‘Manzanillo,’ however, suffered the most immediate and severe defoliation. Defoliation on ‘Manzanillo’ began within 2 weeks after the freeze, usually at the ends of the shoots. On young trees, leaves that remained were located near the bottom of twigs where they join the main branches and trunk. Eventually, usually within 6 weeks, most of these leaves also dropped, and many young ‘Manzanillo’ trees were completely denuded. Older ‘Manzanillo’ trees fared better, but often the entire outer shell of the canopy was defoliated. The ground under damaged trees was often completely covered by leaves. Freeze-damaged leaves could be distinguished easily from leaves dropped because of age. Damaged leaves were still green; aged leaves were a clear yellow as a result of the mobilization of nitrogen from the green chloroplasts to other plant parts before leaf fall.
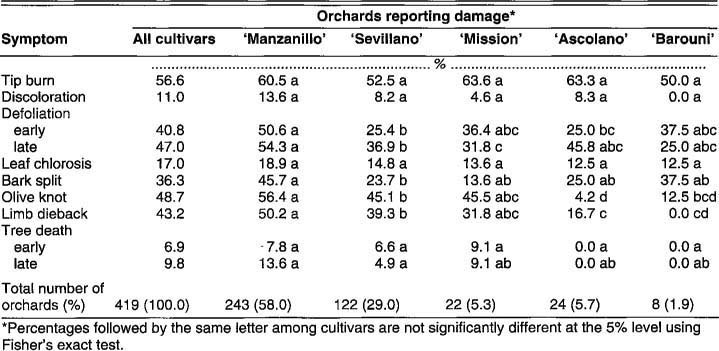

Fig. 4. This young ‘Ascolano’ tree has lost its 2- and 3-year-old leaves prematurely as a result of the freeze. Most 1-year-old leaves have remained on the tree, but many of these show signs of freeze damage nevertheless.
Leaf loss on young ‘Ascolano’ trees developed more slowly. Defoliation, minimal in the first weeks following the freeze, became noticeable by the sixth week. The pattern differed from that seen in ‘Manzanillo.’ On ‘Ascolano,’ mature leaves at the bottom of twigs and stems dropped (fig. 4). Leaves produced during the previous season's growth persisted. Most leaves near the margin of the canopy remained on the trees, except those affected by tip burn. Older ‘Ascolano’ trees were not noticeably defoliated, perhaps because their dense canopies protected the older leaves. Defoliation on ‘Sevillano’ and ‘Mission’ was similar to that on ‘Ascolano.’ Defoliation on ‘Barouni’ was minimal.


Fig. 6. ‘Ascolano’ leaves produced in 1990 and affected by freeze chlorosis and necrosis (top) and leaves produced in 1991 after the freeze (bottom). The 1991 leaves are mostly normal; the one chlorotic leaf may have been damaged by the freeze as a leaf primordium.
Leaf chlorosis
Chlorosis, the loss of green color in leaves, can be part of a natural process, as in aging. Often, however, it signals stress, such as a mineral deficiency affecting the green pigment chlorophyll. The December 1990 freeze caused a spotty or mottled type of chlorosis to develop (fig 5 and fig 6). The upper surfaces of leaves were raised and rough where there was chlorosis. Internally, there were empty spaces caused by the death and splitting apart of cells in the area of the vascular bundles in the leaf's center (fig. 7). Chlorosis was first noticed in ‘Ascolano’ trees and later in ‘Manzanillo’ and other cultivars. Although many chlorotic leaves dropped during the year, others persisted through the end of the growing season and into the winter of 1991–1992. Freezes of considerably less severity than that of 1990 may adversely affect leaves and the production of sugars and other substances necessary for normal tree and fruit growth and development, even though other tree parts have not been damaged.

Fig. 7. Cross-section of a freeze-damaged ‘Ascolano’ leaf. Freezing has killed many cells containing the green chloroplasts in the area near the vascular bundle. The palisade parenchyma is vertically arrayed at the top of the leaf. Here the spongy parenchyma at the bottom seems to have been damaged most. The vascular bundle can be seen in the center, with xylem on top and phloem on bottom. In other freeze-damaged leaves the entire palisade parenchyma and lower epidermis were destroyed.
Bark split
In bark split, the bark and cambium crack open lengthwise, exposing the mature xylem to the air (fig. 8). The exposure of internal tissue allows disease-causing fungi and bacteria to enter. Bark split was especially common on ‘Manzanillo’ twigs. However, such damage was also seen in young ‘Ascolano’ and ‘Mission’ trees. It is likely that twigs of other cultivars also were affected, but not to the extent seen on ‘Manzanillo.’
Anatomical examination of bark split shows damage to the xylem and to the living tissues external to it. It is likely that bark splits are the result of freezing events in the xylem (fig. 9). As ice forms, it expands, and when the bark and other tissues are not elastic enough to withstand the pressure, cracks form to release the pressure. Insufficient bark elasticity may contribute to lack of coldhardiness in ‘Manzanillo.’
Some bark splits may also be caused by freezing in the phloem. Freezing of phloem tissues apparently was common in the 1990 freeze, even in the absence of bark split. Gaps were present in the phloem of young ‘Manzanillo’ and ‘Ascolano’ trees where ice formation killed cells in trunks from near ground level up to 3 feet (0.9 m) high (fig. 10). Olive tree tissues sustained internal damage even in the absence of recognizable external signs. No evidence of the phloem damage seen in February 1991 was found in ‘Manzanillo’ trees examined the following June.
In our studies, the cambium was not damaged, except where xylem freezing had led to bark split; other authors, however, have described freeze damage in the cambium of deciduous trees. A functioning cambium is essential for healing bark splits and for other forms of recovery following a freeze (fig. 10 and fig. 11).
Olive knot
Olive knot disease is caused by a bacterium, Pseudomonas syringae pv. savastanoi. The bacterium is spread externally by water splashing down from sites of previous infection and, to an unknown extent, systemically through the tree's vascular system. Entering the plant through wounds, the bacteria produce the growth hormone auxin (indole-3-acetic acid), which causes uncontrolled plant cell growth and formation of galls. Cultivars vary in susceptibility. ‘Manzanillo’ is very susceptible; ‘Mission,’ ‘Sevillano,’ and ‘Barouni’ are moderately susceptible; and ‘Ascolano’ is the least susceptible. This ranking is supported by the grower survey data (table 9).

Fig. 8. Bark split on ‘Manzanillo.’ The phloem has been killed and the xylem has been exposed. Notice the line on the xylem, indicating that it has split apart. The black spots are signs of rotting fungi.

Fig. 9. Cross-section of a healed-over bark split on ‘Manzanillo.’ Notice the brown color at the origin of the split in the xylem and the line-up through the damaged tissue showing where the expansion of ice split the tissues apart.
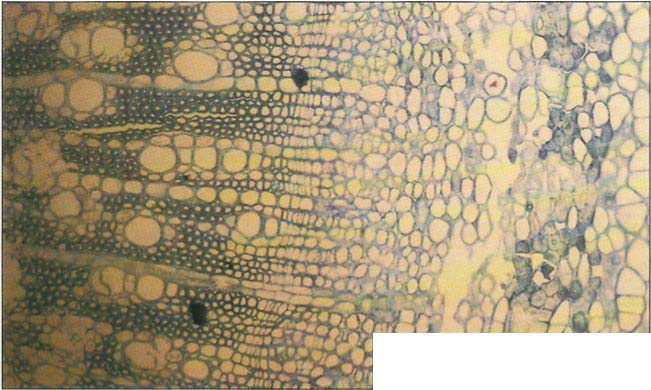
Fig. 10. Below, cross-section of a ‘Manzanillo’ taken 3 weeks after the freeze showing a cleared area where cells in the phloem have been killed by freezing. The xylem cells are the more orderly, round, large and small cells on the left. The cambium is the area of closely packed cells between the xylem and the phloem.
Normally, galls form at leaf scars, pruning cuts, and sites of other physical damage. In addition, when there is freeze damage, twigs, branches, and trunks may develop olive knot (fig. 12). Bacteria may enter many minute cracks in the bark, but studies also indicate that the bacterial infection can develop internally (fig. 13). The disease propagates along the damaged tissue through the length of the stem. Once uncontrolled growth starts, the bark is unable to contain the tumorlike tissue and it splits open. Bark splitting seen later in spring was due to olive knot growth rather than to the mechanical stress caused by expanding ice.
Freeze damage and development of new olive knot was apparent in tissues surrounding the site of previous gall formation. This observation suggested Pseudomonas syringae pv. savastanoi, like other pseudomonads, might be an ice-nucleating bacterium. Such bacteria produce a protein that promotes formation of ice crystals. However, subsequent tests showed that P. syringae is not an ice-nucleating bacterium.
Limb dieback
During the growing season, some large-sized limbs suddenly collapsed and died, leaving brown foliage on branches. Limbs girdled by olive knot and affected by bark splits collapsed once temperatures warmed up.
Deformation of new leaves
Vegetative buds surviving the freeze were not necessarily free of damage. The first few nodes of developing shoots often bore leaves misshapen, twisted, distorted, or cupped. These deformities persisted throughout the season. Later-developing leaves appeared normal.
Damage to fruiting buds
The freeze killed many fruiting buds, even though as a whole olive buds, when not actively growing, are more hardy than other organs and tissues. Damaged branches had fewer inflorescences and these developed more slowly than did those on undamaged branches (fig. 14). Concern was expressed about the viability of flowers that developed from leafless nodes, especially in ‘Manzanillo,’ which had lost much of the leaves produced in the previous year's growth. Despite the concern, many of these inflorescences persisted and were pollinated normally. Except in cases of limb dieback, fruits developed and reached maturity, even without subtending leaves. Overall, however, fruits were smaller than what could be expected normally. Crop maturity was delayed as a result of freeze damage and cooler-than-normal temperatures during the spring and summer of 1991.

Fig. 11. This is a bark split which has healed over with callus tissue produced from the functioning cambium on either side of the split. This kind of healing can occur only if there is no olive knot infection. Notice how the split is directly under a split leaf scar. Freezing may have started in the leaf and propagated to the stem through the vascular system.

Fig. 12. Olive knot may completely infect and kill freeze-damaged tissue. This mature knot was found on a ‘Manzanillo’ tree damaged by a freeze in Glenn County in 1985.

Fig. 13. Olive knot developing internally on a freeze-damaged branch. Eventually the pustule-like centers of cell growth will split the bark open.

Fig. 14. The normal fruiting branch (top) has two inflorescences and two leaves at each node. The three damaged branches (bottom) rarely have two inflorescences at each node. Many of the fruiting buds have been killed, and the inflorescences that remain elongate and develop more slowly in the absence of leaves to nourish them. Chances of a normal crop on the damaged branches are slim.
Economic aspects
The financial impact of the December 1990 freeze is not readily apparent. No adverse financial impact occurred to the olive industry in 1991 on the basis of statewide production and sales. Large crops in 1989 and 1990 provided sufficient crop carryover for 1991 marketing. Present annual marketing commitments require about 80,000 tons [72,560 metric tons (t)] of olives. These marketing plans were based on 1991 crop estimates of from 55,000 to 60,000 tons (48,885 to 54,420 t) of fruit. The actual crop was 63,259 tons (57,376 t), and with the carryover from previous years, the olive industry met marketing commitments with no deleterious effects. However, normal carryover would be insufficient to meet 2 consecutive years of short crops, as may be the case if the 1992 crop was less than 80,000 tons (72,560 t). At the time of publishing this article, we know the 1992 crop will exceed 163,000 tons. By 1996, economists may be able to explain what actually took place in economic terms following the 1990 freeze.
Neither olive cropping patterns nor pricing can be reliably predicted. In the history of the California olive industry only in 1984–1985 and 1989–1990 have there been 2 consecutive years with production greater than 71,000 tons (64,397 t). The usual California olive production pattern is alternate, as is true throughout the olive-growing world. The years of 1989 and 1990, with 119,000 and 126,874 tons (107,933 t and 115,075 t), respectively, were unusual because production was high each year and increased over that of the previous year. Even without the December 1990 cold damage, we expected a great reduction in crop for 1991 following 2 large production years. On several other occasions crops of about 120,000 tons (108,840 t) were followed by crops of 60,000 tons (54,420 t) or less the next year (that is, in 1979, 1981, 1983, and 1987), and this could have been expected for 1991.
In the Sacramento Valley, a few ‘Manzanillo’ orchards were sufficiently damaged to reduce production in 1991. ‘Ascolano,’ ‘Mission,’ and ‘Sevillano’ orchards produced near-normal and more-than-normal crops. In the southern San Joaquin Valley, many ‘Manzanillo’ orchards had insufficient crops in 1991 for commercial harvest. Even in these cases, it is difficult to assess the effect of cold damage on cropping. The 1990 ‘Manzanillo’ crop in that area exceeded 6 tons per acre (13.48 t/ha), and a reduced 1991 crop was expected even without cold damage. As in the Sacramento Valley, ‘Ascolano,’ ‘Mission,’ and ‘Sevillano’ yields were near expectations, despite any cold damage. In some cases, financial losses from a smaller 1991 crop of ‘Manzanillo’ were partly offset by a 17% increase in monetary value per ton.
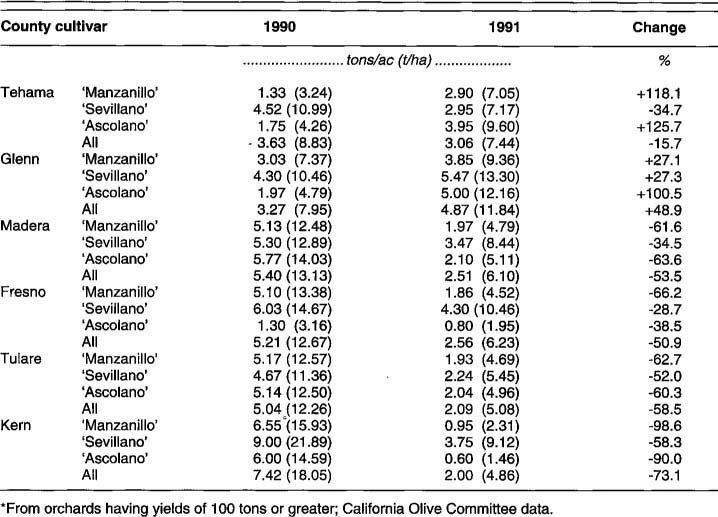
Based on data provided by the COC, yield comparisons were made between 1990 and 1991 for larger producers delivering at least 100 tons of fruit in 1990. Table 10 and Table 11 sum up production from these large producers, about 10% of the state's growers.
Statewide, ‘Manzanillo,’ ‘Sevillano,’ and ‘Ascolano’ showed yield decreases, but ‘Barouni’ showed an increase (table 10). None of the decreases would have been considered exceptional, even without cold damage. No COC data were available for ‘Mission’ because of low production, but data from Butte County, whose major cultivar is ‘Mission,’ showed a decrease in production from 4.5 tons per acre (10.11 t/ ha) in 1990 to 1.7 tons per acre (3.82 t/ha) in 1991. Yield increased in the Sacramento Valley in Glenn County for all varieties and increased for ‘Manzanillo’ and ‘Ascolano’ in Tehama County in 1991 over 1990, but yield decreased in the San Joaquin Valley (Madera, Fresno, Tulare, and Kern counties) (table 11). The striking decrease in Kern County was not unexpected because of high yields in 1990. Nevertheless, both the high 1990 yields and subsequent cold damage probably contributed to reduced 1991 yields. In contrast, the Sacramento Valley had moderate yields in 1990 and returned with good yields in 1991, despite cold damage. This pattern may be explained partly by the dominant cultivars found in the two regions. San Joaquin Valley olive production relies almost entirely on ‘Manzanillo,’ which was most damaged by the freeze and is the most prone to alternate bearing. Production in the Sacramento Valley, on the other hand, relies on cultivars that are more cold-hardy but these cultivars also bear alternately.
Although monetary returns from production are the most important consideration when dealing with economic impacts, other costs contribute to the overall picture. Pruning costs increase if large leader branches in cold-damaged trees continue to die and if olive knot outbreaks occur. Additional fertilization and irrigation to overcome damage in trees may add marginally to the overall economic impact. Putting dollar figures on these aspects is almost impossible at present and will prove difficult in the future. Tree replacement is, however, a readily identifiable cost because this question was included on the grower questionnaire. Thirty-three growers of the 311 respondents to the questionnaire had to replace 3,677 trees; 68.6% were ‘Manzanillo,’ 19.3%, ‘Sevillano,’ 8.1%, ‘Mission,’ and 4.1%, ‘Ascolano.’ No replacement ‘Barouni’ were reported. Average cost per tree was $3.68; total cost was nearly $13,500. If these figures are representative, we speculate that $50,000 may have been spent for replacement trees industrywide. Cost of labor for tree removal and planting could easily reach $30,000. A possible total cost of $80,000 for tree replacements, as a result of the freeze, may represent only a fraction of overall replacement costs to olive producers.

Recommendations to growers
This study has produced the following recommendations to prepare olive growers for the next severe freeze. Most are simply sound horticultural practice.
Conclusions
The Sacramento Valley's olive-growing areas experience temperatures that can damage olive trees with surprising regularity. Near Willows in Glenn County, these freezes may occur every 5 years. In 1990, the lowest temperature recorded in olive-growing areas was 11°F (-11.7°C) near Willows. The occurrence of such freezes is much less in the San Joaquin Valley's olive-growing areas, about once every 17 years at Visalia, for example.

Both field studies and laboratory tests confirm that ‘Manzanillo’ is the least cold-hardy of California cultivars. ‘Ascolano’ is intermediate in coldhardiness. ‘Barouni,’ ‘Mission,’ and ‘Sevillano’ are the most cold-hardy. If fully acclimated, trees of these three cultivars may resist tree death down to temperatures of about 0°F (-17.8°C). Coldhardiness increases with tree age.
Laboratory experiments clearly demonstrate that leaves and wood of olive trees acclimate with exposure to cold temperatures in fall and winter and deacclimate as temperatures warm in spring. The depth of acclimation depends on the cultivar. In contrast, olive tree bark does not appear to acclimate.
Of a variety of damage symptoms seen in the field, those most indicative of damage were defoliation, bark split, and limb dieback. A particular kind of leaf chlorosis caused by freeze damage has been described. Cultural practices such as pruning, irrigation, and fertilization can influence the extent of freeze damage of all cultivars and their rate of recovery.
Olive knot infestations were common on freeze-damaged tissue. However, the presence of olive knot did not necessarily increase the extent of freeze damage, as tests confirmed that the olive knot bacterium is not ice-nucleating. Field studies and grower-supplied information confirm that ‘Manzanillo’ is the most susceptible to olive knot disease and ‘Ascolano’ is the most resistant. ‘Barouni’ is close to ‘Ascolano’ in its resistance; ‘Mission’ and ‘Sevillano’ are intermediate in resistance.
The freeze killed many fruiting buds, and many fruit that were set developed slowly or poorly in the absence of leaves. Olive yields declined in 1991 but largely because of the alternate-bearing habit of the species. Overall economic impact as a result of the freeze is unclear.
Finally, for scientists and growers alike, perhaps the most lasting impression of the 1990 freeze was not the damage to olive trees caused by the freeze, but rather the recovery and regrowth of damaged trees once spring arrived. Clearly, olive trees are remarkably resilient, even after the kind of severe environmental stress that was experienced in December 1990.
Special Section Insert: Lessons from a record-breaking freeze: Some olives show damage; many, coldhardiness

