


All Issues
Genetic improvement of beneficial insect…: Guthion-resistant parasite ready for implementation in walnuts
Publication Information
California Agriculture 45(5):29-31.
Published September 01, 1991
PDF | Citation | Permissions
Abstract
Because pesticides may depress populations of beneficial insects as well as target pests, biological control has historically played a limited role in integrated pest management (IPM). However, recent research has shown that pesticide-resistant parasites selected in the laboratory can be established in the field and enhance IPM programs. Continuing studies indicate that a Guthion-resistant strain selected at UCB performs well in two California orchards, and could be used to colonize other walnut orchards throughout the state. Additional releases would accelerate this process. Otherwise, large-scale establishment might take 5 to 10 years. Pesticide-resistant beneficials — whether selected In the laboratory or genetically engineered — may someday play an expanded role in IPM programs and the reduction of pesticide use.
Full text

Genetically selected for resistance to Guthion, the adult wasp (Trioxys pallidus) is shown attacking walnut aphids.
Genetic selection for resistance to pesticides has been proposed as a method for enhancing the use of parasites in integrated pest management (IPM) programs. However, genetic improvement of parasites is an experimental tactic in biological control, and a series of questions needs to be resolved. First, is it possible to select parasites for resistance to pesticides? The July-August 1988 issue of California Agriculture reported the successful laboratory selection of a strain of the walnut aphid parasite, Trioxys pallidus Haliday, for resistance to Guthion (azinphosmethyl).
Second, is the resistance obtained sufficiently high that the parasite can survive rates of pesticides used in IPM programs? The Guthion-resistant strain of T. pallidus was evaluated for its ability to survive on walnut foliage treated with field rates of Guthion, and the results were reported in California Agriculture, September-October 1989. The results indicated that selection for Guthion resistance in T. pallidus yielded usable levels of resistance. In addition, the strain is cross resistant to Lorsban (chlorpyrifos), Thiodan (endosulfan), and Supracide (methidathion), pesticides used in the walnut IPM program.
Third, will the new strain be effective in the field? The Guthion-resistant strain was released into five walnut orchards in 1988 and the results of the first season were reported in the September-October 1989 issue of California Agriculture. The laboratory-selected strain (1) established in four of the five orchards, (2) survived field rates of Guthion application, (3) parasitized aphids, and (4) dispersed to nearby nonrelease sites.
During the past 2 yr we continued studying this new strain to determine whether, and how, it should be released into California walnut orchards for large-scale implementation. If this new strain could be widely established, growers could use Guthion to control the codling moth, Cydia pomonella (L.), or navel orangeworm, Amyelois transitella (Walker), without disrupting the biological control of the walnut aphid, Chromaphis juglandicola Kaltenbach.
Before attempting large-scale establishment of the Guthion-resistant strain of T. pallidus, we needed additional information on mode of inheritance of Guthion resistance, stability of resistance, and overwintering ability of the resistant strain. Because we assumed that this new strain would have to replace the wild strain in walnut-orchards, we developed a population genetics simulation model to test various options for large-scale releases of this new strain. Results of our experiments will be described, including findings from a release into a walnut orchard near Gridley during 1989. Evidence for overwintering at one of the original 1988 release sites is presented. These results are discussed in terms of possible implementation tactics.
Mode of inheritance
Tests to determine the mode of inheritance of Guthion resistance were conducted using time response tests. Male and female F1 progeny were obtained through mass crosses of the resistant and susceptible colonies. F2 progeny were obtained by allowing F1 individuals to mate with each other. Mortality after exposure to 50 and 0 ppm Guthion was recorded separately for males and females. The time responses of the reciprocal F1 female progeny were similar (data not shown), which indicates that no maternal effects are involved. LT50 (time to 50% mortality) values of the F1 females indicate Guthion resistance is inherited as a semidominant factor (data not shown). Because the F1 parasites are close to the resistant strain in their survival rates, establishment of resistant parasites in pesticide-treated orchards will be accelerated. The results of the time response tests for male and female F2 progeny indicate that Guthion resistance in T. pallidus is determined by more than one major gene.
Evaluating stability of resistance
Between February and August 1990, we conducted a test to determine how long Guthion resistance will persist in mixed populations of resistant and susceptible parasites held without exposure to pesticides in the laboratory. If there are no deleterious effects associated with Guthion resistance, then resistance levels should remain stable in mixed populations. To test this, we mixed males and females of the resistant and base (susceptible) colonies and held them in three cages without selection with pesticides for approximately 7 months. This time interval is equal to approximately 13 generations, which would be equivalent to approximately two field seasons.
Two cages each were set up for the base and resistant colonies as controls. Every other generation approximately 100 to 200 newly emerged parasites were removed from each cage and screened with 50 ppm Guthion to estimate the stability of resistance in the unselected resistant, base, and mixed colonies. In addition, a resistant colony that was selected with Guthion every other generation was tested as another control. Parasites were placed in treated plastic cups with mesh lids, held at 77°F under continuous light and scored after 24 and 48 hr.
Survival of the selected resistant colony (fig. 1, line 1) remained high throughout the experiment, averaging about 98%. The two resistant colonies that received no pesticide treatments also maintained a high level of resistance (line 2) over 13 generations, indicating that the resistance in the pure colonies is stable over a time interval equal to nearly two field seasons. The reason for the drop in survival in April in lines 2,4, and 5 is unknown, but probably was due to experimental error.
The three colonies derived from mixing equal numbers of base and resistant parasites showed declining levels of tolerance to Guthion (fig. 1, line 4) after approximately 7 months without selection with Guthion. We suspect that if the resistant strain interbred with susceptible wild parasites, and the resultant hybrid progeny managed to remain untreated by any pesticides, resistance to Guthion would decline in the hybrid parasites. However, if the hybrids were selected with Guthion (or perhaps with Lorsban, Supracide, or Thiodan), we expect that Guthion resistance would rebound. Line 3 of fig. 1 indicates that survival of a subpopulation subjected to selection with Guthion in generations 6 and 8 increased dramatically.
This experiment suggests that resistance levels in the resistant strain would be maintained for at least 2 yr without periodic selection. However, interbreeding with the susceptible wild population could result in a decline in resistance if selection were not maintained periodically. We do not know how often selection with a pesticide would be required to maintain high levels of Guthion resistance. We can not determine from this experiment whether the reduced resistance is associated with Guthion resistance genes, or to deleterious effects from genetic bottlenecks and inbreeding during laboratory rearing, or to a combination of these factors. However, it is likely that the resistant strain, once released into California walnut orchards, would be selected for its ability to survive under field conditions and that any deleterious effects from laboratory rearing and selection would be reduced.

Fig. 1 Stability of Guthion resistance in T. pallidus colonies held without selection for 13 generations. Colonies were tested with 50 ppm Guthion every other generation. Line 1 shows survival of a control resistant strain periodically selected with Guthion; 2 shows mean (+ or -S.D.) survival of two colonies of the resistant strain not selected with Guthion for 13 generations; 4 shows mean survival of three colonies obtained by mixing equal numbers of resistant and base (susceptible) parasites and holding them without selection, and 5 shows mean survival of the susceptible base colony. Line 3 shows the rapid response of a mixed colony to selection with Guthion in generations 6 and 8.
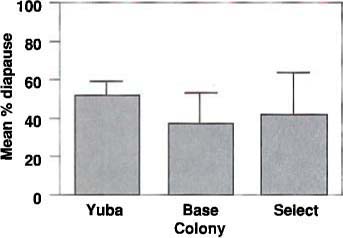
Fig. 2 Mean percentage diapause in the Guthion-resistant (select) strain of T. pallidus compared to the susceptible base and wild Yuba colonies.
Diapause in the resistant strain
Establishment of the Guthion-resistant strain of T. pallidus requires that it overwinter successfully in walnut orchards. During laboratory rearing and selection for resistance, the colonies were maintained under long daylengths and controlled temperatures to avoid diapause induction. The Guthion-resistant strain was reared in the laboratory for several years without determining whether it could diapause. To determine whether it could diapause, we compared diapause incidence in the base and resistant colonies to that of a wild colony collected from Sutter County during June 1990 and reared for only four generations before testing.
Approximately 150 parasites of each colony were used to parasitize aphids on each of two walnut trees held for a week previously at 59° to 61°F under an 8-hr daylength. Parasites were allowed to parasitize the aphids for 24 hr at 77°F under a 24-hr daylength and then removed. The parasitized aphids were then held at 59° to 61°F under an 8-hr daylength for 28 to 30 days. After mummies were produced, they were removed from the foliage and held under these inductive conditions to determine the percentage of each colony entering diapause. The results (fig. 2) indicate that although there are statistically significant differences in diapause incidence among the three colonies (Chi Square = 105, p<0.001), these differences are probably not important biologically. We conclude that the Guthion-resistant strain of T. pallidus is able to enter diapause despite being reared continuously for several years.
Overwintering in walnut orchards
The most important test of fitness and overwintering for the Guthion-resistant strain is whether it can establish, overwinter, and persist in walnut orchards. During 1990, we monitored two walnut orchards where the resistant strain was released. Parasites were released into the Hanford orchard, consisting of Vina and Chico variety walnuts, during 1988 and into the Gridley orchard, consisting of Ashley variety walnuts, during 1989. Parasites were collected from the two orchards, colonized, and tested with 50 ppm Guthion. Survival was compared to that of parasites from the resistant laboratory colony (selected) and two susceptible colonies (base and Yolo).
Hanford
Parasites were collected from the Hanford orchard after each of three applications of Guthion during 1990. Laboratory bioassays were conducted to estimate resistance levels of the three colonies. The percentage survival of the colony that was collected in May after the first Guthion application (HI) in April was significantly higher than that of the Yolo (Y) and base (B) susceptible colonies and nearly as high as that of the selected (S) laboratory colony (fig. 3). These results were repeated with the two colonies collected from this orchard after the second (H2) and third (H3) applications of Guthion. Because the three Hanford colonies were collected extensively outside the release area, and because the resistant strain was released into only 2.5 ac of the 65-ac orchard in 1988, the data suggest the resistant strain has successfully survived two winters, survived repeated Guthion applications, and dispersed throughout the entire Hanford orchard.

Fig. 3. Resistance levels in three colonies (H1, H2, H3) of T. pallidus collected from the Hanford walnut orchard after three applications of Guthion during 1990, compared to the susceptible Yolo (Y), base (B) and selected (S) colonies.
It is likely that strong selection exerted during the 1990 field season, when the Hanford orchard received three applications of Guthion, has enabled the resistant strain to increase in numbers. Unfortunately, we did not monitor aphid and parasite populations from this orchard during the 1990 field season, so we can not indicate whether the resistant strain controlled the aphids. However, the fact that the grower did not treat this orchard for aphids during the 1990 season, despite three applications of Guthion for codling moth, suggests that this strain had an impact on the aphid population.
Gridley
The results obtained from the Gridley block confirm the ability of the Guthion-resistant strain to overwinter. Survival of the colonies collected after the first Lorsban application in 1990 was significantly higher (84%) than that of the Yolo (4.5%) and base (7%) susceptible colonies, and nearly as high as that of the resistant laboratory colony (99%). These results were repeated with the second colony collected from this orchard after the application of Lorsban and an application of Guthion plus Omite (propargite), when 76% of the parasites collected from the orchard survived compared to 5%, 4%, and 98% of the Yolo, base and laboratory resistant colonies, respectively. These data suggest the resistant strain has successfully survived the 1989-90 winter in the Gridley orchard and survived applications of Lorsban, Guthion, and Omite during the 1990 field season.
Model for releases
A computer simulation was used to estimate the effects of biological and operational parameters on introducing and establishing the Guthion-resistant strain of T. pallidus. The simulation model included information on the life history of T. pallidus, responses of the parasites to pesticides, and mode of inheritance of the Guthion resistance. We varied several factors to determine their potential importance, including (1) the initial frequencies of the resistant strain in the release block, (2) population size within the block, (3) selection intensity, (4) refuge size (amount of orchard not treated with pesticide), (5) relative fitness of the resistant and susceptible parasites, and (6) dispersal within and between orchards.
Implementation was simulated as a two-stage procedure. Stage I simulations examined the outcome of releasing the resistant strain into a single 80-ac orchard. Stage II simulations examined the dispersal of the resistant strain from the 80-ac site and its establishment in a matrix of 80-ac orchards in an 8,000-ac region.
The model indicated that establishment within an 8,000-ac region was promoted by migration of the parasites between orchards, low survival of the wild strain when treated with pesticides, and minimal refuge from insecticide applications. Establishment was defined as the resistant strain making up 50% of the individuals in at least 90% of the orchards.
Several factors had a significant negative impact on the rate of establishment of the resistant strain. Under current cultural practices in California walnut orchards, establishment of the resistant strain in the 8,000-ac region was estimated to require at least 5 yr and possibly up to 10 yr. The long time required to replace the susceptible strain with the Guthion-resistant strain was unexpected given that the resistant strain survived 66 times better after a Guthion treatment and 6 times better after an alternative pesticide (such as Lorsban or Supracide) than the susceptible strain in the simulations.
The model simulations suggest that refuge size and total parasite population density are important factors influencing establishment because survival by susceptible parasites in even small untreated refuges can translate into large absolute numbers. For example, 0.1% survival of a susceptible parasite population consisting of 10 million individuals still leaves 10,000 following selection with Guthion. With a refuge size of 10%, one million susceptible parasites would remain following selection with a pesticide. Another important factor is the high reproductive rate of the parasite, which allows susceptible parasite populations to recover rapidly after pesticide applications. Finally, the model indicates that use of pesticides other than Guthion for codling moth control will delay establishment of the resistant strain because susceptible parasite populations recover more rapidly after alternative pesticides than they do after Guthion treatments. Although alternative pesticides select for the resistant strain, the residues are toxic for a shorter interval and result in reduced selection intensity.
Conclusions
The Guthion-resistant strain has apparently overwintered and persisted in the Hanford walnut orchard during 1988-89 and 1989-90 and in the Gridley orchard during 1989-90. The frequency of the resistant strain in these two sites is high, indicating that the strain is well established in these orchards. The Guthion-resistant strain may also be present in other release sites, but we did not evaluate this during 1990. These orchards could serve as a source of resistant parasites that will eventually colonize other walnut orchards throughout the walnut growing region, although the model suggests that large-scale establishment may require 5 to 10 yr if additional releases are not made.
Regional establishment of the resistant strain wilt occur sooner if additional releases are made. Although a mass rearing method has been developed, resistant parasites are not available due to the lack of facilities for mass production. Large scale releases will depend upon whether a rearing program is undertaken by commercial producers of biological control agents or a cooperative facility developed by walnut growers.
Large-scale releases of the Guthion-resistant strain of T. pallidus should be considered for several reasons. First, establishment of this Guthion-resistant strain in more orchards would reduce pesticide applications to control aphids. Second, this new strain offers an opportunity to learn how to implement a genetically-manipulated parasite. Third, Guthion may continue to be used for control of codling moth. Although Guthion may be lost due to resistance in the codling moth or loss of registration, this is impossible to predict with certainty. Furthermore, because this strain is also cross resistant to pesticides other than Guthion, this new parasite strain offers a useful option for walnut IPM.
M. A. Hoy is Professor, F. E. Cave is Staff Research Associate II, and M. A. Caprio is Postgraduate Research Entomologist, Department of Entomology, University of California, Berkeley.
Genetic improvement of beneficial insect…: Guthion-resistant parasite ready for implementation in walnuts

