


All Issues
Tobacco budworm, pest of petunias, can be managed with Bt
Publication Information
California Agriculture 46(4):7-9.
Published July 01, 1992
PDF | Citation | Permissions
Abstract
Damage to petunias by tobacco budworm has reduced the popularity of this colorful summer annual in some parts of California. However, properly timed applications of Bacillus thuringiensis (Bt) can effectively manage budworm in home gardens and in greenhouses. Lacewing applications proved ineffective on petunias, but may control tobacco budworm on other plants.
Full text

The adult tobacco budworm moth.
Although petunias produce abundant summer color, their popularity among home gardeners, landscape professionals, and the nursery industry in and around Sacramento has declined lately because of increasing damage caused by the tobacco budworm, Heliothis virescens (F.). In the Sacramento area, home gardening is widespread, and petunias have until recent times been among the most popular flowers planted. They have also been planted extensively in commercial landscapes such as shopping centers.
Tobacco budworm is a New World species recorded on more than 100 host-plant species including tobacco, cotton, tomato, sunflower, and soybean. Budworm larvae feed principally on reproductive structures (i.e., buds) and growing points. Tobacco budworm was first observed attacking cotton in Southern California in the early 1970s; however, it had been recorded much earlier on ornamentals, such as petunia, geranium, and snapdragon.
In the Sacramento area, budworm moths deposit their eggs on flower buds of geranium in late spring to early summer. Once hatched, the early instars burrow inside the buds; later instars feed primarily on the floral parts, particularly the mature flowers. High densities of larvae of any stage can devour all buds and flowers.
On petunia, the moth usually deposits her eggs on the leaves, and larvae feed on flower buds and opened flowers. Once they have eaten the flowers on a plant, the larvae readily feed on developing seed pods and leaves. The frass occurring on budworm-infested geraniums and petunias is considered unattractive by many home gardeners, as are the late instars crawling or feeding on plants. Although budworms can cause considerable damage to geraniums, they can devastate petunias because the plants are often small and are usually planted as annuals; geraniums, on the other hand, are long-lived perennials that can tolerate budworm feeding pressure from year to year.
We studied the tobacco budworm's impact on petunias in home gardens and in greenhouses (1) to characterize feeding damage to buds and flowers and (2) to identify a safe, convenient, and effective control that could be useful to nurseries and home gardeners.

First-instar green lacewing larvae are entrapped by the sticky exudate from trichomes on a petunia flower, and are thus unable to prey upon tobacco budworm. The larva at left died about 24 hours after entrapment.
Home garden study
Methods. Ten home gardens in Sacramento County were selected for study. (One was ultimately eliminated because of excessive snail damage to its petunias.) Mixed cultivars of hybrid petunias were planted March 14,1988, in split plots of eight plants each (n = 16 plants per site) separated by a distance of at least 4 meters (m). The principal author inspected the plants every 7 to 10 days from May 13 to August 6, looking for evidence of bud-worm eggs or larvae. Geranium plants at all sites were also checked for eggs and larvae. The numbers of damaged and undamaged buds and flowers per plant, along with the numbers of early and late instar larvae per plant, were recorded. The number of plants with visible frass was averaged for each plot, and the amount of snail and slug damage per plant was rated from 0 (no damage) to 20 (maximum damage), converted according to a pre-trans-formed scale, and averaged for each plot.
We evaluated two control tactics. Approximately 10 early instar larvae of a green lacewing, Chrysoperla rufilabris (obtained from Rincon Vitova Insectaries, Inc., Oak View, California), were released per plant at one plot per site on July 5. On July 23, a solution of Bt (Burgess Dipel Bacillus thuringiensis var. kurstaki) was applied via a polyethylene compressed-air sprayer. The Bt was applied at a rate of 15.6 ml/L (about 12 teaspoons/gallon) and sprayed until runoff. Applications were made to plants in one plot per site, but these plots did not necessarily correspond with the plots used for the release of lacewing larvae, since the lacewings had by then proven to be ineffective.
For the purposes of statistical analysis, plots were grouped into two treatments based on applying Bt. Treatments were compared using two-tailed, paired t-tests ($$alpha$$ = 0.05). The response variables we evaluated included the average number of damaged and undamaged buds and flowers, average number of early and late in-star larvae counted on the plants, and number of plants covered with frass. The amounts of snail and slug damage were also compared.
Results. Eggs and larvae of tobacco budworm were first noted on geraniums at one site on May 24. The first sighting of larvae on petunia plants occurred June 14, and by June 21 larvae were observed on geraniums at five sites and on petunias at one site. Petunias at all nine sites were infested by July 5. Production of buds and flowers declined during July, possibly because of slower growth by the plants after peak flowering in June. Damage to buds began in late June and continued throughout July at a constant level, affecting less than 5% of all buds.
Damage to mature flowers was first recorded in mid-June and remained consistently high during July until the end of the sampling period. The proportion of mature flowers per plant that were damaged by budworm feeding often reached 50%. The overall mean density of larvae was highest on July 5, with 20% of all plants infested (although damage was observed on other plants). The number of larvae observed may represent a conservative estimate, because many of the budworm larvae may have been inactive during the day or hidden within floral structures.
No significant differences were found between treatments for frass-covered plants; noticeable frass on plants peaked at 31.1% of plants per combined plot on June 28, and dropped to 9.4% by July 16. The presence of plants with frass did not necessarily indicate feeding damage, since it was possible for the frass to remain on plants well after larvae were gone and a new crop of undamaged flowers had opened. Damage from snails and slugs remained relatively constant among plots and sampling dates during the sampling period, and we found no significant differences.
Lacewings had not significantly reduced the average number of damaged I flowers (t = 0.432, P = 0.906) or budworm i. larvae per plant (t ≈ 0; P > 0.999) in treated t versus untreated plots 4 days after their release. Laboratory observations showed that movement by C. rufilabris larvae was impeded by the sticky exudate of the glandular trichomes that cover petunia leaves; once entrapped or slowed, some of the larvae were actually eaten by budworm larvae. Under laboratory conditions, however, we observed that C. rufilabris larvae had no difficulty preying on budworm larvae on host plants without glandular trichomes. Another contributing factor may have been the switch in environment; C. rufilabris, a native of the eastern United States, is better adapted to high relative humidities than to the hot, dry summers characteristic of the Sacramento area.
The plot to be treated with Bt at each site was randomly assigned, regardless of the previous lacewing releases, since those releases had not statistically reduced floral damage or budworm larvae. Bt was applied 18 days after lacewing release. Significantly fewer budworm larvae (t = 2.57, P = 0.033) were observed on plants treated with Bt 14 days after treatment (fig. 1). Before the Bt application, budworm densities in the plots at each site were comparable.
Greenhouse study
Methods. Petunia plants obtained from a wholesale nursery in Sacramento County in September, 1988, were planted in 6-inch plastic pots and grown in a greenhouse on the University of California campus at Davis. Plants were arranged in a “semirandomized” block design consisting of four treatments (20 plants each): (1) no tobacco budworm larvae; (2) udworm larvae, unsprayed; (3) budworm larvae, sprayed with Bt once; and (4) budworm larvae, sprayed with Bt twice. The treatment without larvae was also sprayed twice with Bt as a precaution and was isolated from the other treatments.
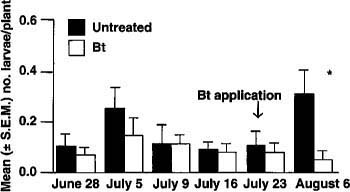
Fig. 1. 1988 tobacco budworm population trends on petunia in Sacramento. The asterisk (*) denotes a significant reduction in larvae following a Bacillus thuringiensis (Bt) application.
The three treatments with budworm larvae were randomized, and the position of the entire noninfested treatment was changed to a different location within the greenhouse at each sampling date. Infested plants within each treatment were arranged to prevent the movement of budworm larvae between plants and among treatments. Sampling began November 1 (when two laboratory-reared larvae were applied per plant to treatments 2, 3, and 4) and ended December 16. Bt was applied November 13 to treatments 1 and 4, and November 13 and 24 to treatments 1, 3, and 4 using the same Bt brand, rate, and type of sprayer as described in the home garden study.
Sampling consisted of counting the number of damaged and undamaged buds and opened flowers for each plant, and noting the presence of frass. We then compared damage levels among the four treatments on a given date using a oneway analysis of variance (ANOVA), followed by the protected least significant difference test for separation of means. The average numbers of undamaged buds and flowers and the numbers of plants covered with frass were compared. The average numbers of undamaged flowers were compared rather than the average numbers of damaged flowers or of budworm larvae per plant (as in the home garden study) for three reasons: petunias grew much better in the greenhouse where they were not overwatered and had full sun, and where flowers did not suffer feeding damage from slugs and snails; the number of undamaged flowers per plant indicated the plant's response to budworm feeding as efficiently as the number of damaged flowers; and two budworm larvae were placed on each infested greenhouse plant, so counting them on the plants would have been difficult, and possibly damaging to the plant.
Results. The mean numbers of undamaged buds did not differ significantly among treatments until 2 weeks after inoculation with worms (November 15), when unsprayed plants had the fewest undamaged buds and uninoculated plants had the most. For the remainder of the experiment, plants that had been sprayed only once had significantly fewer undamaged buds than those that were sprayed twice. A similar response was seen for opened flowers: the first significant differences were noticed November 15, when uninoculated plants and those sprayed twice (first sprayed on November 13) had the most undamaged flowers (fig. 2). This trend continued to the end of the experiment, when twice-sprayed plants had significantly more undamaged flowers than did once-sprayed plants. By the last sampling date (December 16), numbers of undamaged flowers for the twice-sprayed plants did not differ significantly from those for the uninoculated plants. The once-sprayed plants were apparently treated too late to prevent excessive damage by budworm feeding; by the time Bt was applied, most of those larvae were late instars.
Conclusions
Bacillus thuringiensis (Bt) effectively manages tobacco budworm in home gardens and in greenhouses if it is applied when eggs or damage are first noticed and according to the manufacturer's recommended rate. Although Bt was applied to garden plants beyond the peak flowering time, the significant reduction in damage seen in a short time suggests that Bt would be a convenient remedy for home gardeners who notice damage at any time during the flowering season. Because Bt is easy to apply, plants can be treated whenever re-infestations are noticed. Unfortunately, the lag time before Bt kills budworm larvae may vex gardeners who prefer a rapid knockdown; however, budworm feeding should be suppressed shortly after application. Because later budworm instars or severe infestations may be more difficult to control with Bt, some additional damage to flowers may be observed after treatment. Home gardeners should be encouraged to watch any geraniums in their gardens for budworm damage in order to anticipate eventual infestations of petunias, and to apply Bt when they first notice budworms on petunias. A possible benefit of minor feeding by budworms that we observed in both greenhouse and garden sites is a “pruning” of older flowers that may encourage production of new flower buds.
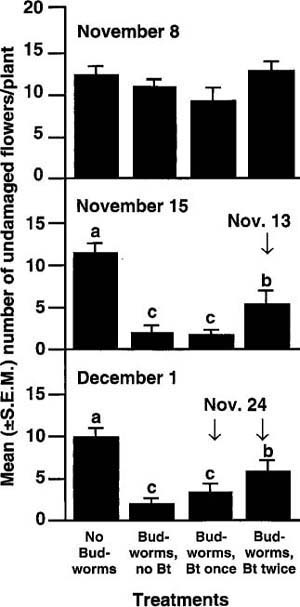
Fig. 2. Tobacco budworm damage to petunia flowers as influenced by applications of Bacillus thuringiensis (Bt), in a greenhouse experiment. Histograms with the same letter are not significantly different at the 5% level.
Budworm feeding damage may be secondary in importance to following proper cultural practices and controlling snails and slugs. Excessive shading, poor drainage, and overwatering may weaken petunia plants and encourage pest attacks. Snails and slugs may be attracted to overwatered plants or those located where soil drainage is poor. Petunias are also susceptible to infection by some soil fungi. Gardeners who observe symptoms of these problems may incorrectly implicate budworms.
In the urban environment, the need for alternatives to traditional chemical insecticides is greater than ever. Commercially available biological control agents have great potential in this market, but considerable research is required to make them a reality. The ineffectiveness of lacewing larvae for budworm control on petunias in the home garden study is a good indication of the need for additional research on implementing dependable biological controls in the urban environment.
Tobacco budworm, pest of petunias, can be managed with Bt

