


All Issues
Vineyard nutrient needs vary with rootstocks and soils
Publication Information
California Agriculture 62(4):202-207. https://doi.org/10.3733/ca.v062n04p202
Published October 01, 2008
PDF | Citation | Permissions
Abstract
Sustainable vineyard fertilization can lead to cost savings while protecting the environment. However, appropriate fertilization conditions depend on the rootstocks, which differ in their uptake of macro- and micronutrients, as well as on the vineyard soils' physical and chemical characteristics, which affect the soil nutrient reservoir. We studied identical sets of 14 rootstocks on three different soils. Rootstocks had a significant impact on petiole levels of nitrogen and potassium throughout the growing season. Pruning weight and fruit yield also varied considerably by rootstock and site. However, rootstock performance was not consistent among sites, nor was the seasonal pattern of change in nitrogen and potassium consistent among sites. The observed differences emphasize the impact of soil texture and nutrient availability on plant growth. Further studies will help guide the development of site-specific sustainable fertilization regimens.
Full text
The fundamentals of nitrogen and potassium nutrition in grapevines are well known. Excess nitrogen leads to high vigor, increasing fruit yield and affecting juice composition (i.e., pH and concentrations of organic acids and esters), but may also create conditions favorable to disease such as bunch stem necrosis and Botrytis cinerea bunch rot (Keller et al. 2001). Potassium deficiency adversely affects ripeness, but excess berry potassium is detrimental to wine quality (Mpelasoka et al. 2003).
While adjusting nutrient input to attain the desired wine quality, viticulturists must also heed the call for sustainable management practices that minimize impacts on soil microorganisms and nutrient balances. The development and application of site-specific fertilization plans can increase sustainability by reducing nutrient runoff into waterways. By developing a better understanding of soil-vine interactions as well as the specific nutrient needs of particular rootstocks and cultivars, we hope to establish site-specific fertilization plans to save money and limit fertilizer input, ultimately promoting sustainability.

In the Sacramento Delta, a vineyard grows in Egbert and Tinnin soil series on a flat alluvial plain.
California vineyards are planted in diverse geographic settings and climates, on soil types ranging from acid to alkaline, fine textured to coarse, deep to shallow, level to sloping, and fertile to less fertile. Several-dozen rootstocks were developed in response to the inadvertent importation into Europe of the grapevine pest phylloxera, from its native eastern North America (Pongracz 1983). The European grape Vitis vinifera is highly susceptible to phylloxera, but many American native species are not. As a solution, the practice of grafting European scions (the grafted fruit-bearing part of the plant) onto phylloxera-resistant rootstocks was developed (Pongracz 1983). This practice is still in use today, and these rootstocks are suited to a variety of conditions reflecting the original environments of the parent plants (Granett et al. 2001). For example, high-vigor rootstocks are used with low-vigor scions on less fertile soils, while low-vigor rootstocks are used with high-vigor scions on fertile soils (Pongracz 1983). Rootstocks also differ significantly in their resistance to drought (Carbonneau 1985).

An Amador County vineyard is planted on rolling hills with Sierra soil series over granitic bedrock.
The range of available rootstocks represents an important resource for the viticulture industry with respect to the long-term sustainability of grapegrowing in California. However, much remains to be learned in order to fine tune the use of these genetic resources in the wide range of California growing conditions. Our current understanding of rootstock nutrient requirements is general yet incomplete, based in most cases on empirical findings.
Nutrient availability, uptake
Soil texture and structure have an important impact on nutrient availability to the plant. Soils rich in organic matter are generally high in available nutrients, including zinc and iron. Clay soils can fix potassium in soil, thereby decreasing the availability of this nutrient to the plant. Rapid leaching can drain nutrients from sandy soils.
Within the root zone, the availability of moisture and its movement in the soil can have significant effects on nutrient availability. Excess leaching may cause nitrogen loss to the water table, and waterlogging may cause denitrification (the conversion of nitrate to nitrogen gas, which occurs where oxygen is in short supply).
Rootstocks also have a pronounced influence on the mineral nutrition of the scion, which should be considered when developing fertilization programs (Koblet et al. 1996). Some root-stocks, such as Malègue 44–53 (44–53), have a higher affinity for potassium than magnesium and therefore may fail to take up sufficient magnesium from the soil. This is compounded by the fact that high levels of potassium in the soil solution can limit the solubilization of magnesium, reducing the availability of magnesium to the plant. (Brancadoro et al. 1994). Other root-stocks, such as Paulsen 1103 (1103P), easily absorb magnesium (Scienza et al. 1986). In high-potassium soils, selecting a “magnesium-absorbing” rootstock may be the easiest way to correct for a deficiency of this nutrient (Brancadoro et al. 1994). Our understanding of rootstock-scion interactions is further complicated by the fact that grape cultivars respond differently to nutrients. For example, Chardonnay and Cabernet Sauvignon have high requirements for magnesium, which can result in deficiencies in this mineral (Loue and Boulay 1984).
Vines grown on different rootstocks may also differ in their tolerance to lime (calcium carbonate) and susceptibility to iron deficiency. High lime content induces chlorosis (a condition in which leaves produce insufficient chlorophyll due to iron deficiency) by slowing iron uptake and translocation (Bavaresco et al. 1992). In calcium-rich soils, total leaf chlorophyll and iron content were higher in Chardonnay grafted onto lime-tolerant rootstocks such as Ruggeri 140 (140R) or Selection Oppenheim 4 (SO4) than on the less lime-tolerant rootstock Millardet et De Grasset 101–14 (101–14) (Bavaresco et al. 1992). Under high salinity conditions, Syrah grafted on Ramsey and 1103P (both salt-tolerant rootstocks) had higher wine potassium, pH and color than on its own roots (Walker et al. 2000, 2002).
Assessing rootstocks and soils
Ideally, vineyard management strategies should consider the site-specific properties of individual soils, the individual requirements of the rootstock and the scion, as well as the relationship between the two. By considering these factors individually and collectively, we will be able to better understand the soil-vine relationship and begin to develop site-specific, sustainable, vineyard management plans.
In this study, we examined the nutrient status and growth characteristics of 14 common rootstocks on three distinct soil types. Two vineyards were located in the Sacramento River Delta near the town of Hood; the scion was Chardonnay on Egbert clay (sandy loam variant) soils at one vineyard, and Cabernet Sauvignon on Tinnin loamy sand soils at the other (Anamosa 1998). A third vineyard was in Amador County's Shenandoah Valley, and the scion was Zinfandel on a Sierra sandy loam soil.
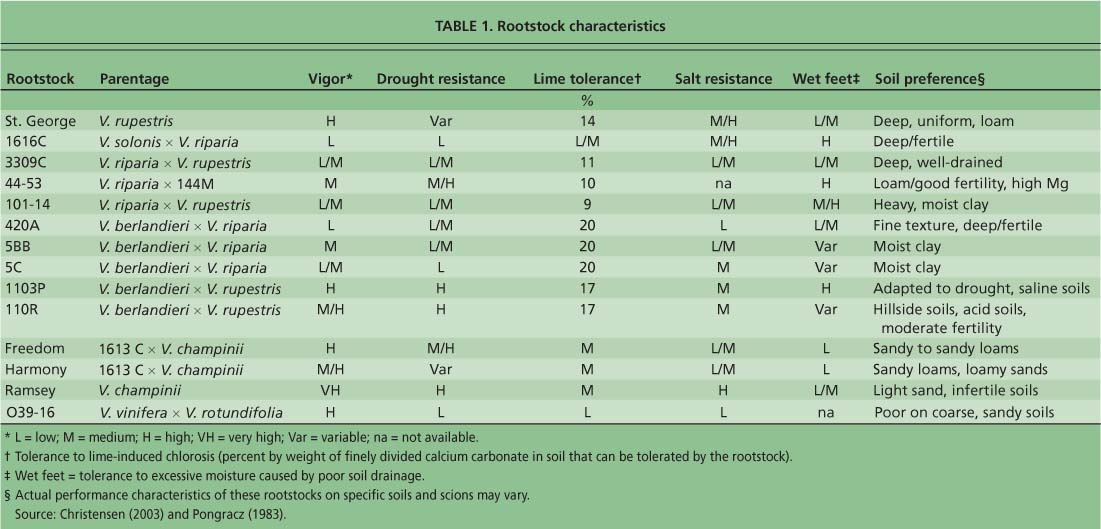
At all three vineyards, we evaluated an identical set of 14 rootstocks: Teleki 5C (5C), Kober 5BB (5BB), Couderc 3309 (3309C), Millardet et De Grasset 101–14 (101–14), Richter 110 (110R), Paulsen 1103 (1103P), Millardet et De Grasset 420A (420A), Couderc 1616 (1616C), Rupestris St. George or Rupestris du Lot (St. George), Malègue 44–53 (44–53), Ramsey, Harmony, Freedom and VR O39–16 (O39–16) (table 1). Twenty-five replicate vines were planted for each rootstock/scion pair. All three sites were drip-irrigated and managed according to routine pest and nutrient management practices. Weeds were controlled by a combination of contact and pre-emergent herbicides, and resident vegetation was present between rows. Vineyards planted with multiple rootstocks were managed uniformly.
Petiole nitrogen and potassium.
The sites were not deficient in nitrogen, and rootstock was the only treatment. At all sites, petiole (leaf stalk) and blade (leaf body) tissues were collected at bloom, veraison (color change at ripening) and harvest. Bloom samples were leaves opposite the basal-most grape cluster. Samples were collected over three sequential years. At each sampling date, 20 petioles and blades were collected per treatment replicate. The petioles and blades were separated, oven-dried and sent to UC Davis for processing and analysis. All samples were analyzed for nitrate-nitrogen, expressed as parts per million (ppm); total nitrogen, expressed as percent nitrogen; and percent potassium. Due to space considerations, only bloom petiole samples will be discussed in detail here.

Fig. 1 Soil textural triangle for three vineyards, showing percentages of sand, silt and clay for each soil horizon. Numbers correspond to horizons, increasing with depth.
![(A) Cation exchange capacity (CEC, cmol[+]/kg), electrical conductivity (EC, deciSiemens per meter [dS/m]) and pH, and (B) exchangeable cations (Ca, Mg, K and Na; meq, milli-equivalents per 100 grams of soil) for each horizon of three vineyards.](http://ucanr.edu/sites/calagjournal/archive/?image=fig6204p204b.jpg)
Fig. 2 (A) Cation exchange capacity (CEC, cmol[+]/kg), electrical conductivity (EC, deciSiemens per meter [dS/m]) and pH, and (B) exchangeable cations (Ca, Mg, K and Na; meq, milli-equivalents per 100 grams of soil) for each horizon of three vineyards.
Soil sampling was performed at each site using a backhoe to dig a sampling pit to a maximum depth of 70 inches. Soil morphology was described as outlined in the U.S. Soil Survey Manual (Soil Survey Division Staff 1993), and samples were collected from all horizons (distinct soil layers).
Geographic location was measured by a Garmin 45XL geographic positioning system. Soil samples were air-dried, ground, sieved to pass through a 2-millimeter grid, and submitted to the ANR Analytical Laboratory for analysis. Gravel content was calculated from the weight of material retained by the sieve. Soil pH was measured in a saturated paste, and electrical conductivity was measured in the saturated paste extract (Sparks 1994). Exchangeable cations (calcium, magnesium, potassium and sodium) were extracted using ammonium acetate, pH 7.0. Sand, silt and clay were measured using the hydrometer suspension method (Klute 1986). Official soil series descriptions were collected from the Web site of the USDA National Soil Series Description Facility in Lincoln, Neb.
Delta Chardonnay vineyard
Soil characteristics.The soils at the Delta Chardonnay site belonged to the Egbert clay loam series, which has subsoil textures of clay loam and silty clay loam. This was the heaviest textured soil of the three studied, ranging from 13% to 50% clay (fig. 1). The cations studied were potassium, sodium, calcium and magnesium. This soil had a fairly high cation exchange capacity (CEC), ranging from 33 to 48 cmol(+)/kg (centimoles of charge per kilogram of soil), and the highest exchangeable cation (EC) content of the three soils studied (figs. 2A, 2B).
Potassium availability was measured by the potassium-to-CEC ratio, which was below the predicted value of 2.5 found in the literature for this soil texture (Champagnol 1984; Etourneaud and Loue 1986) (fig. 3A). The concentration of soluble salts in the soil solution was measured by electrical conductivity, which exceeded 2.5 in the two deepest horizons (fig. 2A). Electrical conductivity values above 2.5 may limit vine vigor (Nicholas 2004). The exchangeable sodium percentage (ESP), defined as the sodium-to-CEC ratio, approached but did not exceed 6, the level above which sodium content can negatively affect vine vigor (Nicholas 2004) (fig. 3B).
Similarly, the calcium-to-magnesium exchangeable cation ratio fell below 1-to-1 in the root zone at soil depths from 14 to 48 inches (excess magnesium is detrimental so the 1-to-1 ratio is a threshold not to be exceeded) (fig. 3C). High soil magnesium can induce potassium deficiency, which negatively affects vine growth and crop load. High magnesium also reduces soil aggregate stability, reducing water infiltration (Dontsova and Norton 2001).
Rootstock performance.In this vineyard, four rootstocks gave above-average fruit yields and pruning weights (the weight of vine canes removed at pruning, a measure of plant vigor): 1103P, 101-14, 1616C and Freedom (fig. 4A). Rootstocks with below-average pruning weights and fruit yield in this soil included 420A, 44–53 and O39–16 (fig. 4A). Rootstock 420A is sensitive to potassium deficiency (Pongracz 1983) and so may have been affected by the lower-than-expected potassium availability for a heavy-textured soil (fig. 3A) (Etourneaud and Loue 1986).
Plant mineral content.Petiole nitrate-nitrogen was on average lower at bloom and veraison, but higher at harvest (386 parts per million [ppm] and 382 ppm versus 947 ppm, respectively). In general, the highest bloom petiole nitrate values were seen in rootstock 1103P, and the lowest in 1616C, 44–53 and Harmony (fig. 4A). Linear regression analysis revealed a significant correlation between yield and petiole nitrate-nitrogen at bloom (r2 = 0.438).
Petiole potassium was higher at bloom and veraison, while much lower at harvest (2.81% and 2.73% versus 1.59%, respectively). The highest bloom petiole potassium values included root-stocks 44–53 and 1616C (fig. 4A). Notably, rootstock 44–53 had the highest petiole potassium levels at bloom in both the Chardonnay and Cabernet vineyards, as well as the third-highest levels in the Zinfandel vineyard (fig. 4).
Delta Cabernet Sauvignon vineyard
Soil characteristics.The alluvial soils at the Delta Cabernet Sauvignon site were mapped as Tinnin loamy sand. The soil at the sampling site was characterized by a loamy surface horizon, and light-textured subsoil horizons that increased in sand content with depth (fig. 1). This soil had the highest pH range of those studied, from neutral at the surface to alkaline in the subsoil (fig. 2A). It also had lower electrical conductivity and exchangeable cation levels than the Delta Chardonnay vineyard, with a relatively low CEC of 10 cmol[+]/kg except at the surface (fig. 2A).
The potassium-to-CEC ratio was in the satisfactory range in the upper horizons, but there was a slight potassium deficiency in the lower root zone (fig. 3A) (Etourneaud and Loue 1986). Sodium content in this soil was low (fig. 3B) (Nicholas 2004). The calcium-to-magnesium ratio was below 1 in the subsoil, indicating a relative excess of magnesium (fig. 3C) (Champagnol 1984).
Rootstock performance.Pruning weights were above average in this vineyard for rootstocks Ramsey, 110R and 1103P, while O39–16 gave the highest fruit yield (fig. 4B). Rootstock 44–53 had the lowest pruning weight, and among the lowest fruit yields.
Plant mineral content.Petiole nitrate-nitrogen declined significantly between bloom and veraison (from 567 to 307 ppm), but by harvest returned to a level similar to that at bloom (data not shown). Rootstocks with the highest petiole nitrate-nitrogen at bloom included Ramsey and O39–16 (fig. 4B).
Petiole potassium levels declined sharply from bloom to veraison and harvest (2.33% versus 1.21%, and 0.38%, respectively). The highest levels at bloom were in rootstocks 44–53 and Freedom, while the lowest were in 420A and 110R (fig. 4B).

Fig. 3 Soil elemental ratios for (A) K/CEC ratio (orange lines indicate recommended values for a given texture class); (B) exchangeable sodium percentage (ESP) (orange line indicates threshold ESP value of 6%, above which salt concentrations may adversely affect vines); and (C) Ca/Mg ratio (values below 1 indicate excess Mg, which may be detrimental to vines).
Amador Zinfandel vineyard
Soil characteristics.The soil at the Amador Zinfandel site was mapped as Sierra coarse sandy loam, a light-textured soil with a large sand fraction and low clay content (fig. 1). In addition, this soil had a high coarse-fragment content, as the Sierra soil series developed from a fractured granitic substratum. This soil had a paralithic contact (direct contact with fractured bedrock) with soft, decomposing granite rock at a depth of 30 inches (Anamosa 1998). Vine roots penetrated to 60 inches in rock cracks. Due to its light texture and high coarse-fragment content, this soil's potential water-holding capacity was very low and would make it sensitive to drought if dry-farmed. It also had a slightly acidic pH with respect to the other two profiles studied (fig. 2A) and a relatively low CEC, and therefore, a small nutrient reservoir (figs. 2A, 2B). However, it also had high manganese content (not shown), likely due to the presence of this element in the parent material and the slightly acidic pH.
For light-textured soils, a satisfactory potassium-to-CEC ratio is in the range of 1.5 (Etourneaud and Loue 1986). In this vineyard, the potassium-to-CEC ratio was highest in the topsoil, likely reflecting an excess of potassium due to fertilization (fig. 3A). The potassium-to-CEC ratio was lower in the subsoil, indicating potassium deficiency in the lower horizons (Etourneaud and Loue 1986). This soil also had a high calcium-to-magnesium ratio, and the lowest exchangeable magnesium of the three sites studied (figs. 2B. 3C).
Rootstock performance.Rootstocks with above-average pruning weights on this soil included 5BB, 1103P, 1616C and Freedom (fig. 4C). Rootstocks with high fruit yields included 5BB, 420A, 110R and 1103P. Rootstocks 44–53, 101–14 and 420A gave the lowest pruning weights, while O39–16 gave the lowest average fruit yield (fig. 4C).
Plant mineral content.Petiole nitrate-nitrogen levels declined sharply for all rootstocks from bloom to veraison and harvest (1,317 ppm versus 80 ppm and 102 ppm, respectively) (data not shown). Large differences in bloom nitrate-nitrogen values among rootstocks were seen, with the highest for O39–16 and 5BB, and the lowest for 420A.
On average, petiole potassium levels were unchanged from bloom to veraison but declined significantly by harvest (2.06% and 2.00% versus 0.87%, respectively) (data not shown). Rootstocks with the highest petiole bloom potassium were Freedom, O39–16 and 44–53, and the lowest were 420A and 110R (fig. 4C).
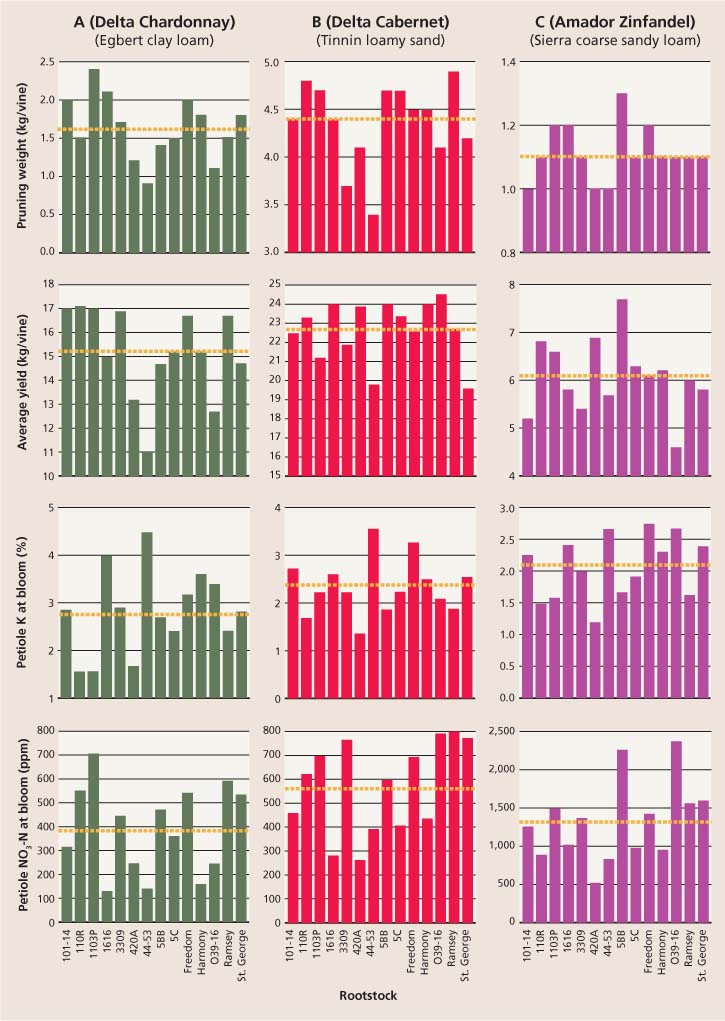
Fig. 4 Yield and plant mineral content at bloom for 14 rootstocks with (A) Delta Chardonnay, (B) Delta Cabernet and (C) Amador Zinfandel. Dashed lines indicate average values for all 14 rootstocks, calculated separately for each vineyard. Values shown are averages for 3 sequential years.
Three sites compared
Rootstocks had an impact on the foliar levels of nitrogen and potassium in petiole tissues at all three sampling dates throughout the growing season. Some rootstocks consistently showed high petiole potassium values in all three vineyards, notably 44–53, which has been previously noted for this characteristic (Champagnol 1984). In contrast, rootstock 420A consistently showed low petiole potassium in all three vineyards. As reported by Wolpert et al. (2005), petiole potassium content at bloom was lower for rootstocks that had Vitis berlandieri genetic backgrounds than for those that did not (fig. 4). In this study, rootstocks with V. berlandieri backgrounds were 420A, 5BB, 5C, 1103P and 110R (table 1).
Pruning weights also varied considerably by rootstock and by vineyard. However, rootstocks differed in their rankings among the three trials. For example, rootstock 5BB had high vigor with the Zinfandel scion, but below-average vigor with the Chardonnay scion; in contrast, rootstock 101–14 had high vigor with the Chardonnay scion but below-average vigor with the Zinfandel scion.
The three soils in this study exhibited large differences in texture, and in physical and chemical properties, which contributed to differences in plant vigor. For example, the Chardonnay vineyard's Egbert clay loam was a heavy-textured soil with high exchangeable cation content. The rootstocks that had the highest pruning weights and fruit yield on this soil were well adapted to clay soils (101–14) and/or humid, fertile soils with moderate salt (1616C).
In contrast to the Egbert clay loam, the Cabernet Sauvignon vineyard's Tinnin loamy sand and the Zinfandel vineyard's Sierra sandy loam were light-textured soils with high sand content. Rootstock 101–14, which had high vigor on Egbert clay, had below-average yield and pruning weight in Sierra sandy loam. Rootstocks Ramsey and 110R had high vigor in Tinnin loamy sand. It should be noted, however, that pruning weight and fruit yields are not the only criteria for vine performance, and other considerations such as berry juice chemistry and sensory characteristics must be taken into account when selecting rootstocks for particular scions.
Despite the site-specific differences in soils, some rootstocks showed similar trends in plant mineral content and vigor at all three sites. For example, rootstock 44–53 had below-average vigor, petiole nitrogen and nitrate-nitrogen at bloom in all three vineyards.
Plant nutrient levels can be influenced by scion-specific differences in nutrient metabolism (Christensen 1984), and scion genotype can also affect root-stock performance (Virgona et al. 2003). In the present study, the variability observed in rootstock performance also suggests a potential role for rootstock-scion interactions.
Tailored vineyard fertilization
Additional trials are needed in the diverse environments in which grapevines are grown within California, in order to better match rootstocks and scions to particular soil types and local edaphic conditions (such as soil water content, pH, aeration and nutrient availability). As we learn more about the nutrient input requirements of specific rootstocks and scions, the measurement of plant nutrient levels, and the physical and chemical properties of soil, site-specific fertilization management programs can be tailored to individual vineyards. The ultimate goal of such programs is to decrease fertilization costs and environmental pollution, thus promoting sustainability. Future studies will include rootstock trials on soils with different physical and chemical properties in an effort to increase our understanding of the soil-vine relationship.
Vineyard nutrient needs vary with rootstocks and soils

