


All Issues
For switchgrass cultivated as biofuel in California, invasiveness limited by several steps
Publication Information
California Agriculture 67(2):96-103. https://doi.org/10.3733/ca.v067n02p96
Published online April 01, 2013
PDF | Citation | Permissions
Abstract
The expected production of biomass-derived liquid fuels in the United States may require cultivation of millions of acres of bioenergy crops, including perennial grasses such as switchgrass. Switchgrass is not native to California and possesses many qualities in common with other perennial grasses that are invasive. To evaluate the potential invasiveness of switchgrass in California, we conducted risk analysis and climate-matching models as well as greenhouse and field evaluations of switchgrass, looking at its environmental tolerance and competitive ability against resident riparian vegetation. We concluded that dryland regions of California are not suitable to vigorous establishment and invasion of switchgrass. However, riparian areas appear to be far more likely to support switchgrass populations. With effective mitigation practices in place throughout the development, growth, harvest, transport and storage processes, it should be possible to minimize or eliminate the movement of seeds and vegetative propagules to sensitive habitats. Consequently, we believe that switchgrass is unlikely to become a significant problem in California, even with widescale production.
Full text
Biomass crops in the United States are projected to yield 136 billion liters of biomass-derived liquid fuels by 2022 (Heaton et al. 2008). The expectation is that this will require cultivation of between 54 and 150 million acres of bioenergy crops. Furthermore, state and federal greenhouse-gas reduction initiatives have incentivized widespread cultivation of biofuel crops. Of the crops under consideration, perennial nonfood grasses are the leading candidates. To be successful in this role, these bioenergy grasses will need to possess many agronomically desirable traits, including broad climatic tolerance, rapid growth rates, high yields, few natural enemies and resistance to periodic or seasonal soil moisture stress (Heaton et al. 2009).

Some characteristics that make for an outstanding biofuel crop will make the same plant a hardy, hard-to-manage weed if it escapes cultivation. Switchgrass can thrive if it escapes into an ecosystem with an abundant, year-round water supply, so it's best grown far from such areas.
One of the leading candidates among bioenergy grasses is switchgrass (Panicum virgatum L.) (Parrish and Fike 2005; Pedroso et al. 2011). Switchgrass is a perennial warm-season (C4) bunchgrass native to most of North America east of the Rocky Mountains, where it was historically a major component of the tall-grass prairie. It was included in the initial screening for biofuel crops in the United States in the 1970s and was determined to be the model bioenergy species by the Department of Energy (McLaughlin and Walsh 1998). This was primarily due to its broad adaptability and genetic variability (Sanderson et al. 2006). Over the past three decades, breeding efforts have developed several cultivars, many of which produce dense stands, tolerate infertile soils and readily regenerate from vegetative fragments (Parrish and Fike 2005). These cultivars are often separated into upland ecotypes (Trailblazer, Cave in Rock, Blackwell and Sunburst) and lowland ecotypes (Alamo and Kanlow) (Pedroso et al. 2011).
Switchgrass is not native to California and was, in fact, included for a brief time on the California Department of Food and Agriculture (CDFA) Noxious Weed List (CINWCC 2007) due to concerns about its potential invasiveness. Although there was one documented report of an escape of switchgrass from cultivation in Orange County, California (Riefner and Boyd 2007), there are no known records of its escaping elsewhere or causing any ecological or economic damage, despite its long-time use as a forage and conservation species (Parrish and Fike 2005). Since its removal from the CDFA Noxious Weed List, it has been the focus of yield trials throughout California (Pedroso et al. 2011). Because of the state's Mediterranean climate, the yield potential is high; however, the crop will require significant water and nitrogen inputs.
Invasive and biofuel grasses
In an ideal system, biofuel crops should be cultivated in a highly managed agricultural setting similar to that of most major food crops, such that the crop could not survive outside of cultivation. Under such conditions, the likelihood of escape and invasion into other managed or natural systems would be very small. Unlike biofuel species, most food crops have been selected for high harvestable fruit or grain yield. This nearly always results in a loss of competitive ability, typically accompanied by an increase in the addition of nutrients and often pesticides.
When a biofuel crop is grown for cellulose-based energy, the harvestable product is the entire aboveground biomass. To be economically competitive, such perennial crops should be highly competitive with other plant species, harbor few pests and diseases, grow and establish rapidly, produce large annual yields and have a broad range of environmental tolerance, while also requiring few inputs per unit area of water, nutrients, pesticides and fossil fuels (Raghu et al. 2006). Few species fit these requirements better than rhizomatous perennial grasses, primarily nonnative species (Barney and DiTomaso 2008; DiTomaso et al. 2007). However, these qualities and traits are nearly identical to those found in harmful invasive species (Raghu et al. 2006). For example, species such as johnsongrass (Sorghum halepense (L.) Pers.) and kudzu (Pueraria montana [Lour.] Merr. var. lobata [Willd.] Maesen & S.M. Almeida) were introduced as livestock forage or for horticultural use but have escaped cultivation to become serious weeds in many areas of the United States. In selecting biofuel crops, a balance must be struck between high productivity with minimal inputs, on the one hand, and risk of establishment and survival outside the cultivated environment (DiTomaso et al. 2007) on the other.
Johnsongrass, like switchgrass, was first cultivated as forage, but it subsequently escaped and has become one of the world's most expensive weeds in terms of control costs (Warwick and Black 1983). It is currently listed as a noxious weed in 19 U.S. states. When comparing switchgrass to johnsongrass and to corn, a typical agronomic grass crop, it is clear that switchgrass possesses many growth traits similar to those of weedy johnsongrass and only a few similar to those of corn (table 1). While this is not direct evidence that switchgrass will be a significant invasive or weedy species, it does suggest that the risk may be greater than for more typical agronomic crops.
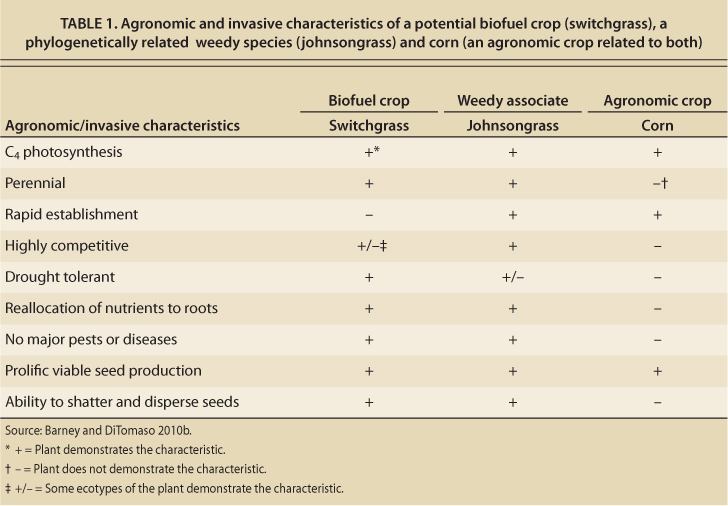
TABLE 1. Agronomic and invasive characteristics of a potential biofuel crop (switchgrass), a phylogenetically related weedy species (johnsongrass) and corn (an agronomic crop related to both)
Although cultivation of switchgrass and other biofuel crop species may ultimately prove a net benefit to society, the environmental risks associated with their potential escape into natural and managed systems should be assessed before the crops are commercialized and introduced into new regions. A pre-introduction evaluation can be performed in tandem with typical agronomic yield trials. This approach can both quantify ecological risk and assess the suitability and economic performance of the species within a particular region.

Traditional crops are bred to enhance production of food or fiber, but biofuel breeders emphasize hardiness and quick, abundant growth—“weedy” characteristics. Here, researchers grow switchgrass in PVC towers, making it easier for them to monitor root growth and the effects of water stress.
To assess the invasive potential of any proposed biofuel species, including switchgrass in California, we propose a seven-step evaluation protocol (table 2) (Barney et al. 2012a). These evaluations should be performed for all candidate genotypes and cultivars as well as for transformed genotypes of native species because ecological interactions can vary widely within a species. Science-based information generated from risk assessments, biofuel crop ecological studies, niche modeling and other evaluations can guide risk mitigation decisions at appropriate points within biofuel research and development, crop selection and production, harvest and transportation, storage site selection and conversion/refinery practices (DiTomaso et al. 2010).

TABLE 2. Protocol for evaluating the degree of risk that a biofuel crop (including cultivars and genotypes) will become invasive in a region
Risk assessment
Risk assessment tools have been used in Australia and New Zealand as an aid in decision making for the proposed introduction of novel species for horticultural, agronomic and other purposes. For potential biofuel species, risk assessment should serve as a basic first step in evaluating their invasive potential, whether the species are exotic, native, novel constructs (e.g., hybrids) or genetically modified.
We performed a risk assessment (Barney and DiTomaso 2008) for switchgrass in California using the Australian model (Pheloung et al. 1999). Our analysis produced an inconclusive result, with an “evaluate further” classification (table 3). The first question in the risk assessment asks whether the species is domesticated. A “yes” response favors acceptance (or reduced risk of invasion), under the assumption that domestication generally reduces the inherent weediness of wild types, which are wild plants not selected for production traits (Pheloung et al. 1999). As previously discussed, this is true for most agronomic and horticultural species, but the opposite is true for biofuel crops because selection (breeding) for favorable biofuel crop characteristics generally enhances “weedy” characteristics (Raghu et al. 2006). When we answered the first question differently, the outcome changed from “evaluate further” to “reject.”
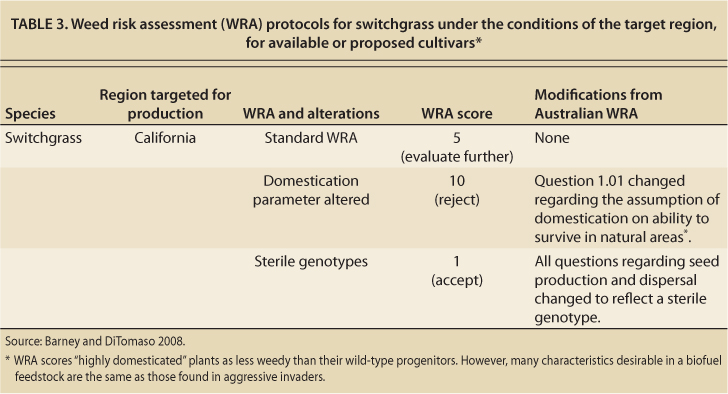
TABLE 3. Weed risk assessment (WRA) protocols for switchgrass under the conditions of the target region, for available or proposed cultivars
We further evaluated switchgrass as a hypothetically sterile cultivar. In this case, the weed risk assessment yielded an acceptably low risk that it would become invasive. This suggests that seed production may be key to the potential invasiveness of switchgrass. However, a lack of seed production does not guarantee a low risk of invasion, considering that the giant reed (Arundo donax L.), which is sterile, is highly invasive in California. The invasiveness of giant reed is due to its ability to regenerate from stem nodes after the stem is detached from the rhizome as a result of flooding or control efforts.
Despite the invasive potential that our analysis shows for switchgrass in California, there are no documented cases of the species escaping in agricultural or natural systems. This, however, may be a function of the limited number of opportunities for introduction of switchgrass propagules (seeds, rhizomes and stem nodes) outside of intentional planting areas. It is also important, therefore, to conduct studies that will quantify switchgrass performance in various ecological settings in order to mitigate the risk of propagule escape and establishment.
Climate-matching analysis
The natural distribution of a species is largely controlled by climate factors, with precipitation and temperature playing the dominant roles (Sutherst 2003). Bioclimatic envelopes, or climate matches, can provide both an estimate of range suitability for the biofuel crop species outside cultivation and the agronomic potential of the biofuel crop in the target region (Barney and DiTomaso 2011). There are numerous methods for estimating the bioclimatic envelope, including CLIMEX, Maxent, GARP, BIOCLIM, classification and regression tree, and simple logistic regression. CLIMEX has been used to model the distribution of biocontrol agents (Poutsma et al. 2008), poikilothermic (cold-blooded) animals (Sutherst et al. 2007) and many invasive plant species (Holt and Boose 2000; Kriticos et al. 2005; Pattison and Mack 2008). The strength of CLIMEX for invasive species applications is that the model can be based on the historical range (e.g., from herbarium data) and supplemented with empirically derived biological and physiological data (Sutherst et al. 1999).
We performed a CLIMEX analysis of switchgrass using the plant's native range as a basis in building the model and then supplementing it with environmental tolerance data from greenhouse studies (discussed below). In a global model of potential suitability, the potential cultivatable range of switchgrass was very broad, both with and without irrigation inputs (Barney and DiTomaso 2010b, 2011; see fig. 1). Subsequent analysis of potential suitable habitat in the western United States (fig. 2A) indicated that much of the region is unsuitable for switchgrass, likely because of the very dry summers of arid and Mediterranean climatic regions (Barney et al. 2012a). However, when adequate yearlong soil moisture was available (i.e., with summer irrigation or a permanent water source), the suitable range of switchgrass increased dramatically throughout much of the western United States (fig. 2B). This could indicate that the successful cultivation of switchgrass would depend upon summer irrigation, while any escape from cultivation and invasion into natural areas would likely be confined to riparian or wetland areas with a permanent water source. Riparian systems are the most heavily invaded habitats in the Central Valley of California, as they possess the primary limiting resource of soil moisture (Levine 2000). Furthermore, riparian areas often border production fields, and traversing them would be unavoidable during biofuel biomass transport.

Fig. 1. Global CLIMEX climate-matching results for switchgrass. The colors represent the CLIMEX ecoclimatic index (EI), where cream (EI ≤ 10) is “unfavorable,” light green (11 ≤ EI ≤ 20) is “suitable,” dark green (21 ≤ EI ≤30) is “favorable,” and blue (EI ≥31) is “very favorable.” The yellow regions are those with an EI > 20 when a permanent water source is available (Barney and DiTomaso 2011).

Fig. 2. CLIMEX climate-matching results for switchgrass based on climate preferences estimated for (A) the nonnative range of the western United States and for (B) the nonnative range of the western United States assuming yearlong access to water (e.g., land along a stream or land that is irrigated). The colors represent CLIMEX ecoclimatic index values (EI; 0–100), where higher numbers represent a more suitable environment (see figure legend) (Barney and DiTomaso 2011).
The CLIMEX model does not forecast yield potential, but it does demonstrate that some regions of California are suitable for establishment and persistence of switchgrass. In support of our suitability prediction in California, Pedroso et al. (2011) evaluated the agronomic potential of switchgrass in four regions of the state. This evaluation showed high productivity in both study locations in the Central Valley, which was considered a highly suitable region based on our bioclimatic index (fig. 2B). Switchgrass grown in the Imperial Valley (El Centro), near the margin of highly suitable climatic conditions in our analysis, also produced high yields. In contrast, yields and survival of switchgrass were lowest in the most northern, cooler, mountainous region of Tulelake, near the Oregon border in northeastern California. Our model also predicted that this region of the state would be poorly suited to switchgrass establishment, even with irrigation.
Environmental tolerance
Each biofuel species should be evaluated for various physiological and environmental tolerances. This information can be used to identify the ecosystems that are most susceptible to invasion and can also be integrated into risk analysis and bioclimatic and agronomic models to estimate, and subsequently mitigate, the likelihood of invasion (Barney and DiTomaso 2008). Based on results from our CLIMEX analysis of switchgrass in the western United States, water availability should be the major limiting factor for switchgrass naturalization. To test this, we conducted a greenhouse study to evaluate switchgrass's tolerance of soil moisture stress at various levels of water availability, ranging from moisture deficit to flooded, and we also assessed the germination, establishment, performance and reproductive potential of four common ecotypes, both upland and lowland (Barney et al. 2009).
Our results showed that cultivars of switchgrass performed well in both well-watered control and flooded conditions. Although switchgrass survived extended periods without water, individual plants in drought treatments were shorter, with lower measurements for leaf area and specific leaf area, and they produced fewer tillers and less biomass (fig. 3). As expected, lowland types outperformed upland types in the flood treatment and also displayed higher fitness under most conditions, which likely explains why they are the target of germplasm improvement for biofuel cultivation (Parrish and Fike 2005).
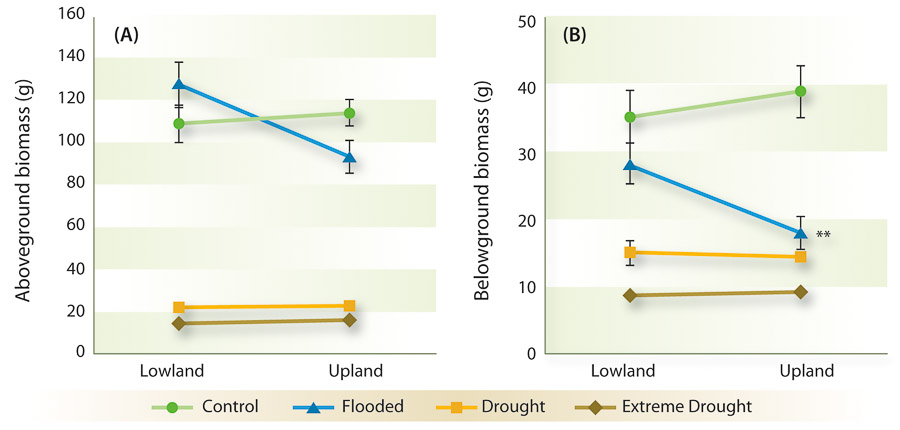
Fig. 3. Aboveground biomass (A) and belowground biomass (B) for lowland and upland switchgrass under control, flooded, drought (5–10% soil moisture) and extreme drought (< 5% soil moisture) soil moisture treatments. Lines followed by double asterisk (∗∗) indicate significant difference (p < 0.05) between upland and lowland cultivars for that treatment (Barney et al. 2009).
We concluded that switchgrass, particularly lowland ecotypes, has the ability to germinate, establish and flower in low moisture and even more so in flooded conditions. The evidence further supports the climate-matching data and indicates that soil moisture is the limiting factor in the establishment and growth of switchgrass in regions of the western United States. While tolerance to a range of soil moisture conditions may increase the cultivatable range of switchgrass, it also suggests that the species is not likely to be very competitive in natural areas exposed to prolonged drought, as is common in much of California.
In another study, we grew switchgrass in outdoor mesocosms (3-meter-tall tubing) under irrigated and rainfed (dry) conditions and assessed the spatial distribution and abundance of roots using minirhizotron images and whole-root-system sampling (Mann et al. 2012). Although plants survived extended periods of drought, their shoot and root biomass, root length density, numbers of culms and culm height were greatly reduced under dry conditions. These data support the results of the greenhouse study (fig. 3). The rainfed treatment reduced switchgrass whole-plant biomass by 83%, culm production by 67% and root length density by 67% from the levels of irrigated plants. However, switchgrass grew roots continuously into regions of available soil moisture as surface soil layers grew increasingly dry (fig. 4). This study suggests that switchgrass is able to tolerate drought by mining deep soil moisture. A deep-rooting habit and continuous root growth from regions of water depletion to moister regions are strategies used for drought avoidance by plants exposed to periodic water stress (Lambers et al. 2008).

Fig. 4. Distribution of switchgrass belowground biomass (means + standard deviations) in rainfed and irrigated conditions. Note the logarithmic scale of the x-axis. Letters indicate significant differences (at 95% confidence level) between depths averaged over both treatments (Mann et al. 2012).
It is important to note that while switchgrass survived dry, rainfed conditions, its performance was significantly reduced. This level of performance would be unacceptable for agronomic production and would also reduce the ability of switchgrass to establish and compete with resident vegetation in drier natural areas.
Invasibility and competitiveness
The results of our climate-matching analyses as well as the biological and physiological studies allowed us to identify habitats that were most susceptible to invasion by switchgrass. From our previous work, we knew that riparian corridors (e.g., streams, irrigation canals) and perhaps even rice production fields are the regions most likely to be susceptible to switchgrass invasion in California. In subsequent work (Barney et al. 2012b), we confirmed these findings by introducing switchgrass propagules into a riparian habitat under controlled conditions and evaluating their colonization, survival and establishment potential under varying levels of soil moisture availability and competition. The results supported our greenhouse and mesocosm studies, again demonstrating that while switchgrass can survive under drought conditions, its performance on upland sites away from streams was very poor compared to that of switchgrass plants adjacent to the stream. This confirms our conclusion that riparian regions of the state are the areas most potentially susceptible to switchgrass invasion, while dryland regions of California have very low susceptibility to invasion. Of equal importance, switchgrass grown without competition in the first year in the wet habitat produced about six times more tillers than switchgrass growing in an intact resident plant community with competition, and the tillers were twice as tall and yielded eight times the aboveground biomass (fig. 5). This further indicates that, even in a suitable habitat, switchgrass is not highly competitive with other vegetation.

Fig. 5. Switchgrass (Panicum virgatum = PAVI) (means ± standard error) aboveground biomass in lowland habitats along Putah Creek. Treatments include switchgrass (PAVI) with competition (PAVI + Comp) and without competition (PAVI – Comp) from resident herbaceous vegetation (Barney et al. 2012b).
Propagule biology
The probability of establishment of an invasive population is directly proportional to the propagule pressure from outside sources (Barney and DiTomaso 2008). In the case of switchgrass, outside sources will be production fields, harvest and transportation equipment and biomass storage sites. Our initial risk assessment for the invasive potential of switchgrass in California determined that seed production and dispersal were the means of the greatest threat that it would become invasive (Barney and DiTomaso 2008). To aid in their efficient conversion into energy, cellulosic biofuel species are typically harvested after senescence in the field, usually in late fall. In our seed biology experiments, we showed that switchgrass germinates and survives under conditions that range from 10% soil moisture to submersion in water (Barney et al. 2009). From these experiments, we estimate that an average switchgrass field would produce between 300 and 900 million seeds per hectare. Using a conservative estimate of 300 million seeds per hectare and 60% dormancy, approximately 120 million seeds per hectare would be capable of germinating, given adequate soil moisture conditions (Barney et al. 2009). This tells us that mitigation practices will be needed to reduce the risk of seeds spreading to sensitive ecosystems. Mitigation practices could include the planting of sterile cultivars, cleaning equipment before moving it to other areas and using closed transport systems and storage facilities.
Hybridization potential
As with genetically modified food and feed crops, screening for possible cross-hybridization with related and desirable species should be obligatory to reduce the chance of genetic contamination or creation of novel hybrids (DiTomaso et al. 2007). In California, there are five native species and five introduced species within the genus Panicum (Baldwin 2012). To date, there is no evidence of hybridization between switchgrass and any other Panicum species, regardless of its native origin. Thus, the likelihood that switchgrass would either contaminate the gene pool of native Panicum species or enhance the weediness of nonnative Panicum species through hybrid vigor seems very small.

Researchers are assessing a number of plants for their potential as biofuels, hoping to find crops that will produce well in a variety of different growing environments. Here, PVC towers planted with giant miscanthus grow alongside towers planted with switchgrass to see which does better in this setting.
Mitigation recommendations
In August 2009, the U.S. Invasive Species Advisory Committee (ISAC), a group of nonfederal experts and stakeholders chartered under the Federal Advisory Committee Act of 1972, adopted nine recommendations for the federal government's biofuel programs (table 4). The recommendations comprehensively address biofuel production and use, as well as the necessity of agency and private sector stakeholder cooperation for effective implementation of the recommendations (DiTomaso et al. 2010).

Initially, all federal agencies with authority relevant to biofuel production should be identified, their likely responsibilities on the invasiveness issue determined, and their ability to minimize the risk of biofuel escape and invasion strengthened as necessary. To reduce the risk of escape, the biofuel crops that are promoted should not be currently invasive or should pose a low risk of becoming invasive in the target region. In addition, biofuel crops should be propagated in production sites that are least likely to impact sensitive habitat or create disturbances that would facilitate invasion. Most importantly, effective mitigation protocols need to be developed to prevent dispersal of plant propagules from sites of production, transportation corridors, storage areas and processing facilities. Minimizing harvest disturbance can also reduce the potential for dispersal and off-site movement of propagules. Prior to widescale planting, multi-year eradication protocols should be developed that are based on integrated pest management (IPM) strategies. Such practices should be readily available, and appropriate information should be distributed with the purchase of biofuel crop seeds. These control methods are not only critical for preventing the dispersal of biofuel crops from abandoned production sites, they are a necessary component of an effective early detection and rapid response (EDRR) system for biofuel crop populations that do escape active management. Throughout this entire process (fig. 6), all stakeholder groups should be engaged, from biofuel development to conversion.

Fig. 6. Simplified biofuel supply chain showing (1) crop development, (2) crop importation and dissemination, (3) crop production, (4) feedstock harvesting, processing, transport, and storage, and (5) feedstock conversion. Black arrows indicate links where propagule or feedstock movement is involved (Barney and DiTomaso 2010b).
In prior publications, we have identified five mitigation points along the biofuel supply chain, or biofuel pathway, in the seed-to-fuel lifecycle of switchgrass or any other biofuel crop (Barney and DiTomaso 2010b):
Breeding or selection programs can target the development of cultivar traits that reduce the risk of escape and invasion. For example, (1) sterile switchgrass cultivars can decrease the likelihood that the plant will escape from production fields. In contrast, (2) enhancement of other environmental traits such as seedling vigor or tolerance to drought or salt may increase the risk of escape from cultivation and invasion into surrounding environments.
(3) Within the production process, growers can reduce the risk of propagule dispersal by cultivating plants away from known dispersal corridors (e.g., streams) or by creating buffers between production areas and transportation corridors. In addition, (4) routine scouting of field margins and bordering habitat for switchgrass escapes, followed by (5) prescriptive management of escaped populations, can help prevent invasion into sensitive habitats.
Seeds present the greatest risk for invasion. Although it may not be economically possible to do so, harvesting before flowering or seed set would reduce the likelihood of propagule production. In addition, the cleaning of planting and harvesting equipment prior to movement from one field to another will help to prevent switchgrass seed dispersal.
Transportation of biomass from grower fields to refineries and storage locations creates another potential opportunity for switchgrass seed dispersal. To mitigate against this, refineries should organize with growers to coordinate efficient harvest and transport practices that minimize propagule loads to outside environments. When applicable, transport from fields should be done with closed trucks or shipping containers.
While switchgrass biomass refineries will presumably operate year-round, stored plant biomass will likely be harvested once, or at most twice, a year—mostly from midsummer through late fall. This means that the biomass will be baled and stored for most of the year either on grower property or, more likely, on refinery property near the conversion facility. These storage sites may serve as seed reservoirs if not managed properly.
Switchgrass invasive potential in California
Based on our evaluation of the potential invasiveness of switchgrass in California, we conclude that dryland regions of California have very high resistance to switchgrass establishment and invasion. However, riparian areas (i.e., streams, irrigation canals, rivers, lakes and ponds) are capable of supporting the establishment of switchgrass. Our data also indicate that the agronomic production of switchgrass in the western United States will require irrigation during the dry season. Most vegetation in the Central Valley of California germinates following late fall or early winter rains to maximize usage of the limited precipitation. Switchgrass, however, germinates or emerges primarily in late spring, toward the end of the rainy season, not long before the rapid drying of the surface soils in summer. These conditions are not conducive to switchgrass establishment and probably account for the poor survival and growth of seedlings in our studies (Barney et al. 2012b). In addition, our climate-matching analysis indicates that the limiting abiotic factor in the northern range of switchgrass in California is cold temperature (Barney and DiTomaso 2010a, 2011). While this may be the current situation, genetic improvement and climate change may expand, contract or shift suitable regions.
Naturalization will be further limited by the competitiveness of resident plant communities (Barney et al. 2012b). However, switchgrass may be able to establish following disturbance in high-resource sites where resident vegetation is removed or damaged, and it may become more competitive following establishment.
Having considered all aspects of our invasive analysis, we conclude that while switchgrass is not without risk of invasion in riparian or wetland areas, it is unlikely to become a significant problem, particularly when mitigation practices are employed to minimize propagule movement. If this crop is developed, grown, harvested, transported and stored responsibly, the ecological risk should be acceptably low and in balance with the need for sustainable energy production.
For switchgrass cultivated as biofuel in California, invasiveness limited by several steps

