


All Issues
Botryosphaeria-related dieback and control investigated in noncoastal California grapevines
Publication Information
California Agriculture 62(4):161-166. https://doi.org/10.3733/ca.v062n04p161
Published October 01, 2008
PDF | Citation | Permissions
Abstract
Dieback, or “dead arm,” in noncoastal California grapevines is most commonly caused by Botryosphaeria spp. Using Koch's postulates, we demonstrated that isolates of B. obtusa are pathogenic on grapevines. We initiated studies to investigate the life cycle of B. obtusa and ways to control it with cultural practices. Fungal spores disseminated by rainstorms were collected in traps in an Arbuckle vineyard from December 2006 through spring 2007. The data suggests that B. obtusa was rain-disseminated throughout winter and spring, and that pycnidia on deadwood in the vines is a major source of inoculum for new infections. Transmission may also be possible via vegetative propagation, pruning shears and insects. Durable latex paints were investigated for protecting pruning and surgical wounds; a self-priming latex paint was shown to be an effective barrier and was nonphytotoxic.
Full text
Trunk and cordon cankers that cause vine dieback are serious economic problems in vines 12 years and older. Vines are infected, at least primarily, through pruning wounds. Historically, dieback in California vineyards was attributed to the fungus Eutypa lata, but many of the vines, particularly in the Sacramento and San Joaquin valleys, are actually infected by fungi in the genus Botryosphaeria (Urbez-Torres et al. 2006); cankers caused by Botryosphaeria spp. are called “Bot canker.”
Signs of disease on grapevines
Between fall 2004 and spring 2007, we monitored ‘Zinfandel’ grapevines in Arbuckle, Calif., in an approximately 18-year-old vineyard with many dead spurs and cordons (arms of a grapevine). Our sampling included the extensive dissection of 36 vines in decline. There were two predominant signs of disease in woody tissue: a brownish, often wedge-shaped necrosis (dead tissue) in cross-sections of cankered regions, and dark brown to blackish streaks in longitudinal sections of wood adjacent to pruning wounds. B. obtusa, identified by a combination of microscopy and DNA sequencing of a portion of the ribosomal DNA (called ITS), was routinely isolated from the margins of cankers in woody tissue in all parts of the vines. B. dothidea, B. stevensii and E. lata were also occasionally isolated.
Except for a loss of vigor in shoots adjacent to cankered regions, the vines were relatively free of foliar symptoms. Nonetheless, in springtime some vines with multiple dead spurs had deformed, chlorotic (yellow) leaves consistent with symptoms of Eutypa dieback. In all 21 vines with foliar symptoms sampled, only B. obtusa was isolated from the margins of discolored woody tissue.

Pycnidia of Botryosphaeria obtusa form on prunings on the vineyard floor. Left, B. obtusa pycnidia are primarily clustered in aggregates (white outline) with some separate individual pycnidia (circle). Right, immature pycnidium (IP) are still partially buried in the plant tissue; mature pycnidium (MP) before spore release; and discharged pycnidium (DP). Microscopic examination of spores is required to identify pycnidia as B. obtusa rather than other Botryosphaeria spp.
B. obtusa pycnidia were observed, generally infrequently and at low density, on the surface of completely dead wood on vines. (Pycnidia are flask-shaped structures that contain conidia, which are asexual spores.) In contrast, pycnidia were observed frequently on prunings and detached wood on the vineyard floor. Individual pycnidia are black and approximately 0.01 inch (0.25 millimeter) in diameter. They are generally aggregated but sometimes separate. When first formed, the pycnidia are submerged in wood, but as they mature they erupt above the trunk surface. After conidia are released during rainy weather and disseminated via wind-blown rain, the empty pycnidial cavities remain on the plant surface. B. obtusa pycnidia were never observed on grape berries, and no sexual spores of either Botryosphaeria spp. or E. lata were ever observed.

In a pathogenicity test, Botryosphaeria obtusa caused a wedge-shaped lesion.
Pathogenicity of B. obtusa
When we started our work, the literature was unclear on whether B. obtusa isolates are pathogenic or saprophytic (living on dead tissue). To test for pathogenicity, own-rooted ‘Cabernet Sauvignon’ with stems approximately 0.7 inch in diameter were inoculated in the greenhouse with a disc of fungal mycelium (a mass of fungal hyphae) into a wound 5 inches above the soil. After 1 year the plants were examined, Koch's postulates were completed and the experiment was repeated with similar results. (Koch's postulates are a process that allows one to conclude that a particular organism causes a particular disease.)
The greenhouse pathogenicity test produced the two major symptoms that we had observed in the field in infected wood: a brownish, wedge-shaped necrosis in a cross-section and brownish to black streaks in a longitudinal section. B. obtusa isolates are pathogenic, and B. obtusa grows more quickly in woody tissue than E. lata (table 1). Our results are in agreement with other reports in which B. obtusa and other Botryosphaeria spp. are identified as important grapevine pathogens (Leavitt and Munnecke 1987; Phillips 1998, 2002; Savocchia et al. 2007; van Niekerk et al. 2004, 2006).

Stands were used to collect fungal spores disseminated in rainstorms. Left, each stand had a funnel under the cordon for estimating available inoculum within the vine, and a plate on the ground (2.5 inches high by 20 inches wide) at a 45° angle to collect splash from debris; right, stands also had two tented plates (8 inches high by 12 inches wide), which were designed to collect wind-blown rain.
Life cycle of B. obtusa in a vineyard
Spore dissemination.
Rain is critical in the life cycle of Botryosphaeria spp. at several stages, including the oozing of conidia out of the pycnidia and then the dissemination of conidia in windblown rain. To monitor spores (the reproductive cells) quantitatively in the vineyard, we used funnels under the vines and constructed plates to collect wind-blown rain. Four replicate spore collectors were placed in each of two fields on Dec. 1, 2006, and removed on May 8, 2007. In order to inhibit germination during collection, the receptacle bottles contained sufficient acetic acid for a final concentration of 12% or more. Rainwater (figs. 1D-G) and prunings on the ground (figs. 1B-C) were collected. After rainwater was filtered through membrane filters with grids, conidia were quantified microscopically at 100X. Weather data were obtained from an on-farm monitoring station and the Nickel's Soil Laboratory in Arbuckle (fig. 1A).

B. obtusa was observed in all rainwater collections; other Botryosphaeria spp. and E. lata were occasionally observed. The most spores were trapped in collectors designed for within-vine drip under an infected cordon in a “no sanitation,” area; that is, with neither surgical removal of infected wood, painting of pruning wounds or burial of prunings (fig. 1E). Conidial dissemination occurred throughout the pruning period and into the springtime (May 8) in 2007, in apparent contrast to winter-only dissemination of E. lata in California.
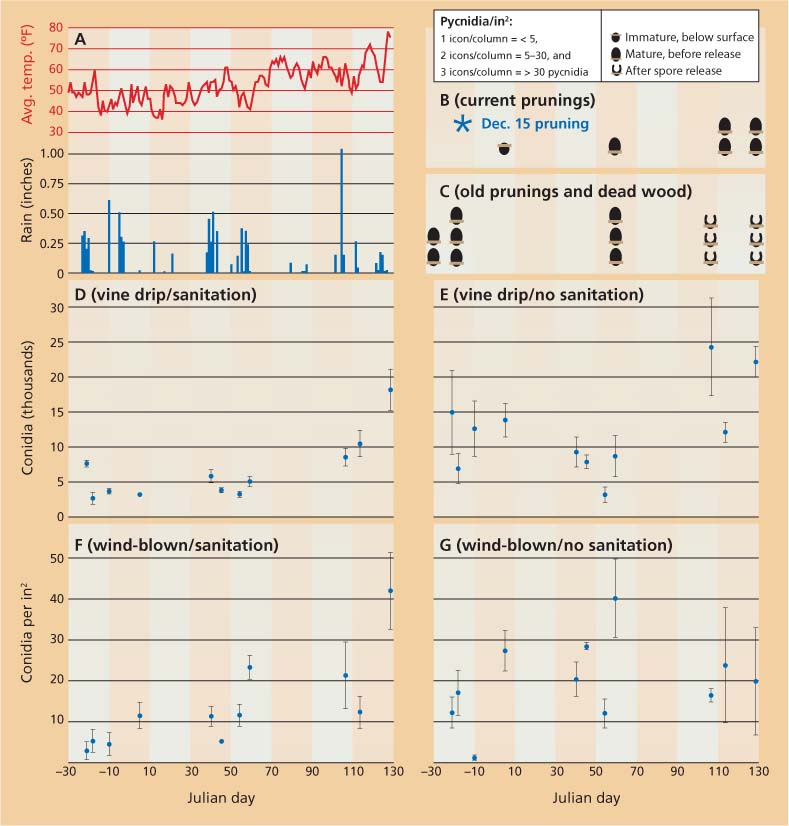
Fig. 1 ▴ Biological and meteorological data from a vineyard in Arbuckle, Calif., between Dec. 2, 2006 (Julian day − 30), and May 8, 2007 (Julian day 128) (Julian dates start with 1 on Jan. 1 of each year). (A) Daily precipitation and average air temperature. (B, C) Development and abundance of pycnidia on (B) current prunings (∗ indicates Dec. 15 pruning); and (C) old prunings and dead wood on the vineyard floor. Prunings were partially submerged in soil and partially exposed; examinations occurred each day that spores were collected. lcons are not shown on examination dates when there were no changes from the previous date. New pycnidia were produced on older prunings and wood in multiple years. (D-G) Means ± SE of rainstorm-disseminated spores retrieved in various collectors in a field trial with (D, F) surgical removal of deadwood in the vines, painting of all surgical and pruning wounds and sanitation of pruning debris on the ground, and (E, G) without these measures. (D, E) Total number of Botryosphaeria obtusa conidia collected in a 7.25-inch-diameter funnel below the vine. (F, G) Wind-blown B. obtusa conidia collected on plates facing north, south, east and west.
Our collection plates for wind-blown rain in the “no sanitation” field (fig. 1G) similarly showed conidial dissemination throughout the December-to-spring observation period. We monitored the development of pycnidia on the current season's prunings from Dec. 15 and on older prunings and wood debris, partly because pycnidia on deadwood in the vine are hard to see and partly because debris on the ground may provide inoculum. Prunings from December 2005 and before, and deadwood on the ground (fig. 1C), primarily released conidia between March 20 and April 14, when it rained on 6 days (fig. 1A). Pycnidia from the current season's prunings matured during 2007, but conidia were not released; release presumably occurred sometime during the 2007–2008 rainy season (fig. 1B). B. obtusa conidia also were collected in angled plates designed to determine if the wind could pick up debris from the soil, although the densities were lower than shown in figs. 1F-G (data not shown).
The sanitation treatment consisted of surgically cutting vines to the trunk, painting surgical and pruning wounds, and burying pruning debris (figs. 1D, 1F). Because there were no pycnidia on these regenerated vines, both the vine drip (fig. 1D) and wind-blown (fig. 1F) are estimates of wind-blown inoculum from outside of the sanitation area. Inoculum concentration in the wind was higher later rather than earlier in the season (fig. 1F, slope of linear regression P = 0.0002).
Infections through wounds.In addition to observing conidial release from pycnidia (fig. 1C) and conidia in all of our traps in springtime (figs. 1D-G), we also had evidence of infections introduced into wounds made when suckers were yanked out of the trunks in springtime. After surgically cutting vines that had at least one infected cordon down to the trunk in December 2004 and allowing them to regenerate for one season, we completely removed 14 vines in December 2005. B. obtusa infections were observed in yanked sucker wounds in 9 of the 14 (64%) vines. Copes and Hendrix (2004) reported faster conidiation of B. obtusa at 64°F and 72°F than at 54°F and 86°F. Given temperatures in California (fig. 1A), spring rains may facilitate the most important B. obtusa infection events.
Wood discoloration, a grapevine response to infection, appears to occur approximately 1 year after fungal invasion. Both the patterns of wood discoloration and the recovery of B. obtusa — from incremental segments of shoots from pruning wounds to the shoot terminus — indicate that B. obtusa primarily infects through pruning wounds. In contrast to E. lata, B. obtusa grows into shoots. In infected vines, we isolated B. obtusa in 28% of asymptomatic shoots and 90% of symptomatic 1-year-old shoots (n = 54) 3 inches from the cordon. Symptomatic shoots that were surface-disinfected and incubated in a humid chamber at 74°F for 3 weeks produced B. obtusa pycnidia. Overall, our data are consistent with a life cycle in which B. obtusa grows asymptomatically within grapevine shoots and woody tissue. We postulate that damage to the vine occurs primarily by the grapevine's release of self-defense compounds that kill its own cambium.

Top, Duration paint is being tested as a treatment for surgical cuts on grapevine trunks; lower, an untreated vine.
Although we postulate that pruning wounds are primarily infected by B. obtusa conidia disseminated in wind-blown rain, some inoculum may also be disseminated by other mechanisms. Pruning cuts through mature pycnidia can release conidia onto shear blades. Even in cases with only mycelium and no spores, transmission is possible via pruning shears. We made pruning cuts on 25 shoots that we knew, retrospectively, were infected. After making the cuts, we wiped the blades onto sterile paper and then cultured the paper; B. obtusa was recovered on 24% of the wipes.
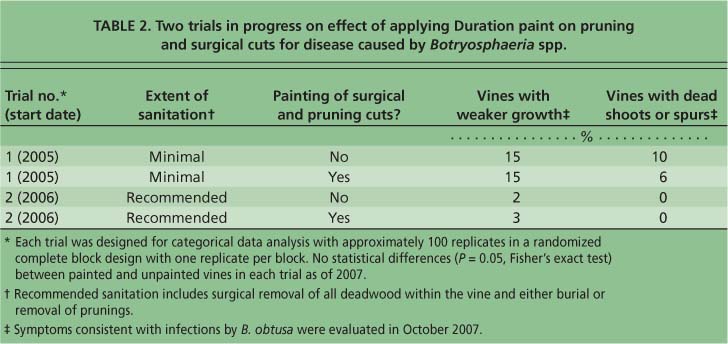
TABLE 2 Two trials in progress on effect of applying Duration paint on pruning and surgical cuts for disease caused by Botryosphaeria spp.
In preliminary experiments, in which duct tape covered with the sticky product Tanglefoot was placed over pruning wounds, we recovered rove beetles (Staphilinidae) infested with B. obtusa. We also detected B. obtusa in material that could have been used for vegetative propagation. As indicated above, we isolated B. obtusa from asymptomatic shoots. B. obtusa also was isolated from the internal tissue of buds that were surface-disinfected, in 13% of the buds (n = 60) of symptomatic shoots and 9% of the buds (n = 57) from asymptomatic shoots on infected vines.
Strategies for cultural control
Surgical removal.Several options are available for surgical treatment: (1) the terminus of the affected cordon(s) can be removed, or (2) most of the scion can be removed, retaining only sufficient wood for the regeneration of new canes (see sidebar). In either case, the retention of a mature root system allows rapid regeneration of the scion. The best location for the surgical cut is not always clear. Certainly, cuts should be made below all necrotic and symptomatic tissue. We have isolated B. obtusa in wood up to 4 inches in front of the discolored margin. Although dieback symptoms often appear to be most severe at the ends of the cordons, multiple infection foci can be distributed across the length of asymptomatic portions of the cordon (data not shown). Consequently, we favor cutting down to the trunk.
However, trunks can be infected too. Of the 36 sampled vines, 28 (78%) had infections on the trunk that emanated from wounds on the trunk; wounds were made either when suckers were yanked out or when the mechanical harvester injured the trunk. Moreover, sampling of the wood just above the surgical cut indicated that 10 of the 36 vines (28%) had infections of B. obtusa at the surgical cut, of which only three had any indication of discoloration. Although in some cases the mechanical harvester appeared to have damaged the vines, we never had evidence that B. obtusa in the trunk per se caused vine debilitation. Based on patterns of discoloration in the trunk, infections of B. obtusa appear to grow primarily toward the base of the stem. While we observed infections in the rootstock, we never observed infections in the roots. A long-term study is needed to determine whether over time, B. obtusa in the trunk leads to a reduction in yield.
Protection with durable paint.We are investigating two strategies for integrated pest management of Bot canker: (1) the protection of surgical wounds and new pruning wounds with a durable paint and (2) the reduction of inoculum sources. Two field trials are in progress to test wound protection (table 2). In 2005, we surveyed paints for use as protectants and selected candidates with the following properties: high elasticity (reduced likelihood for cracking); breathability (presumably less likely to mold under the painted surface); durability to exposure to ultraviolet light and rain; and fungal resistance. After preliminary trials, we selected Duration (Sherwin Williams K34T154) paint, a self-priming exterior latex with an ultradeep base that was stained with 6 ounces of N1 Raw Umber per gallon.
Duration paint forms an impenetrable and stable physical barrier on grapevine surgical cuts and pruning wounds. Experimental treatments in which the paint was applied onto pruning wounds and over buds indicated no phytotoxic or inhibitory effects on foliage or bud out-growth (data not shown). The California Department of Pesticide Regulation stated that Duration paint can be used as a physical barrier on grapevines without fungicide registration. Possible advantages of paint over fungicide include that the paint is extremely long-lasting, will not wash off during repeated rains and is not phytotoxic. Nonetheless, painting is labor-intensive for a large vineyard. We were not successful in delivering Duration paint using a modified pruner due to Duration's high viscosity, particularly under the cooler weather conditions when pruning is typically done.
In collaboration with Sutter Home Vineyards, Zinfandel vines with die-back symptoms — at least one dead spur — were surgically cut on the trunk approximately 12 inches above the soil in February 2005 (table 2). The surgical cuts were either treated with Duration or not. In the paint-treated vines, all pruning cuts were painted. In 2006, an additional trial with surgically treated vines was started in an area with sanitation. Paint coverage was good in both trials, except when there were heavy rains and sap pushed up a new coating of paint. In this case, paint was reapplied. In both trials, new growth and the establishment of new cordons have been excellent. However, we cannot recommend Duration until data on disease incidence and yield are collected in the next several years. Currently, there is no statistical difference between the incidence of vines with B. obtusa symptoms in paint-protected versus nonprotected vines (table 2).
Inoculum reduction.Our and others' data is consistent with the hypothesis that Botryosphaeria spp. may infect vines via multiple avenues. The presence of B. obtusa pycnidia on deadwood on vines and the collection of conidia in funnels below infected wood suggest that the surgical removal of deadwood should reduce inoculum. Because prunings and deadwood on the vineyard floor can produce large quantities of conidia, we recommend ground sanitation, such as the burial or removal of prunings. Further research is required to determine the contribution of this inoculum to vine infections.
B. obtusa may be transmitted by pruning shears. In a study on the potential transmission of grapevine pathogens in infected propagation material, Aroca et al. (2006) detected Botryosphaeria spp. and Phomopsis along the length of some plants, but mainly at the graft union. Consequently, grafting tools may transmit inoculum. Gimenez-Jaime et al. (2006) identified B. obtusa and other Botryosphaeria spp. in grapevine nurseries. Our isolation of B. obtusa in shoots and buds also suggests that B. obtusa might be spread in propagation material.
We monitored rainstorm-disseminated spores over the period when grapevines are pruned, from December to May. Our data indicates that B. obtusa conidia are disseminated during the entire season, including and perhaps especially in spring when temperatures are more conducive for sporulation. Consequently, while later pruning will avoid exposing wounds to early-season rain events and is a worthwhile component of an integrated pest management strategy, it may not provide sufficient control in an infected vineyard.
Research questions
The development of a cost-effective, sustainable Bot canker management program will require additional knowledge. Answers to the following questions will be useful: How long are pruning wounds susceptible to infection by Botryosphaeria spp.? What training and pruning strategies minimize the area of susceptible wounds? How does the timing of conidial release vary in different weather conditions? In different vineyards, how important are various modes of pathogen dissemination, and in particular, are Botryosphaeria spp. initially disseminated in vegetatively propagated material that is infected but not symptomatic, and if so, how often? How long are vines productive after major surgery? And do infections occurring below the surgical cut affect the longevity of the vines?
Surgery is being tested in some vineyards as a means to revitalize vines with Bot canker (see sidebar, see page 164). We postulate that a combination of sanitation and surgery will allow sustainable control of Bot canker.
Botryosphaeria-related dieback and control investigated in noncoastal California grapevines

