


All Issues
As carbon dioxide rises, food quality will decline without careful nitrogen management
Publication Information
California Agriculture 63(2):67-72. https://doi.org/10.3733/ca.v063n02p67
Published April 01, 2009
PDF | Citation | Permissions
Abstract
Rising atmospheric concentrations of carbon dioxide could dramatically influence the performance of crops, but experimental results to date have been highly variable. For example, when C3 plants are grown under carbon dioxide enrichment, productivity increases dramatically at first. But over time, organic nitrogen in the plants decreases and productivity diminishes in soils where nitrate is an important source of this nutrient. We have discovered a phenomenon that provides a relatively simple explanation for the latter responses: in C3 plants, elevated carbon dioxide concentrations inhibit photorespiration, which in turn inhibits shoot nitrate assimilation. Agriculture would benefit from the careful management of nitrogen fertilizers, particularly those that are ammonium based.
Full text
Atmospheric carbon dioxide (CO2) has increased about 35% since 1800 (from 280 to 380 parts per million [ppm]), and computer models predict that it will reach between 530 and 970 ppm by the end of the century (IPCC 2007). This rise in carbon dioxide could potentially be mitigated by crop plants, in which photosynthesis converts atmospheric carbon dioxide into carbohydrates and other organic compounds. The extent of this mitigation remains uncertain, however, due to the complex relationship between carbon and nitrogen metabolism in plants (Finzi et al. 2007; Johnson 2006; Reich et al. 2006).
Carbon metabolism provides the energy and carbon molecules to synthesize organic nitrogen compounds in plants, whereas nitrogen metabolism provides the amino groups for all proteins (fig. 1). Proteins include all enzymes that catalyze (facilitate) biochemical reactions in plants, including carbon metabolism. Any environmental perturbation that interferes with nitrogen metabolism sooner or later inhibits carbon metabolism.

The rise in atmospheric carbon-dioxide levels — about 35% since 1800 — changes how plants metabolize important nutrients, which in turn alters food quality and nutrition, influences where plants and crops can grow, and affects pest management and other cultivation practices. Lesley Randall of the UC Davis Department of Plant Sciences attends to plants growing in hydroponic culture under elevated carbon-dioxide atmospheres, in environmental chambers at the UC Davis Controlled Environment Facility.

Fig. 1. Major processes of the biogeochemical nitrogen cycle. Fluxes (red numbers) are in teragrams (Tg = 1012 g) N/year. Terrestrial organisms and soils contain organic nitrogen that is active in the cycle. Assuming that the amount of atmospheric molecular nitrogen remains constant (inputs = outputs), the mean residence time of nitrogen in organic forms is about 370 years. Source: Bloom 2009.
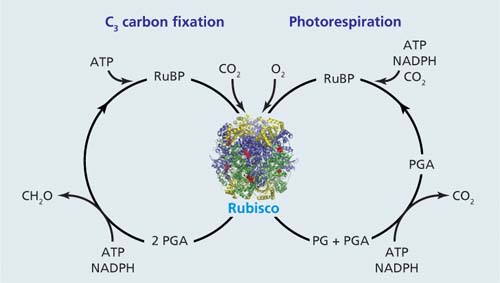
Fig. 2. C3 carbon fixation and photorespiration pathways in which the enzyme rubisco (ribbon model in center) catalyzes reactions between a 5-carbon sugar, RuBP (ribulose-1,5-biphosphate) and either CO2 or O2. The first stable products of C3 carbon fixation are two molecules of PGA (a 3-carbon compound, 3-phosphoglycerate); the first stable products of photorespiration are one molecule of PGA and one molecule of PG (a 2-carbon compound, 2-phosphoglycolate). High-energy compounds ATP and NADPH, generated from photosynthesis, drive these reactions. As atmospheric CO2 increases, there is an initial increase in C3 carbon fixation (and sugar productivity), while photorespiration is inhibited. We have shown that inhibiting photorespiration diminishes nitrate assimilation. In plants that depend on nitrate as a nitrogen source, this eventually inhibits plant productivity and lowers protein content. Nitrogen is part of the amino groups essential to all proteins, and proteins include the enzymes that facilitate biochemical reactions. Source: Bloom 2009.
Carbon dioxide acclimation
The focal point of crop responses to rising carbon dioxide levels is the enzyme rubisco (ribulose-1,5-bisphosphate carboxylase/oxygenase). Rubisco is the most prevalent protein on Earth and contains as much as half of the nitrogen in plant leaves. It catalyzes two different chemical reactions: one reaction combines a 5-carbon sugar RuBP (ribulose-1,5-bisphosphate) with carbon dioxide, and the other reaction combines this same sugar with oxygen.
The reaction of RuBP with carbon dioxide produces a 6-carbon compound that immediately divides into two molecules of a 3-carbon compound (3-phosphoglycerate), hence the name C3 carbon fixation (fig. 2). These products pass through an elaborate biochemical cycle (Calvin-Benson cycle) that eventually forms one molecule of a 6-carbon sugar (fructose-6-phosphate) and regenerates RuBP.
The reaction of RuBP with oxygen oxidizes the RuBP, splits it into one molecule of a 3-carbon compound (3-phosphoglycerate) and one molecule of a 2-carbon compound (2-phosphoglycolate), and subsequently releases carbon dioxide, hence the names C2 pathway or, more commonly, photorespiration. In total, photorespiration consumes biochemical energy, but does not result in any net production of sugar (Foyer et al. 2009). Thus, photorespiration has been viewed as a wasteful process, a vestige of the high carbon dioxide atmospheres (over 1,000 ppm) under which plants evolved (Wingler et al. 2000).
![Net carbon dioxide assimilation (photosynthesis) as a function of carbon dioxide concentrations within a leaf (Ci) for C3 plants grown at either ambient (365 parts per million [ppm]) or elevated (567 ppm) atmospheric carbon dioxide concentrations, in free air CO2 enrichment (FACE) plots, where plants growing in soil under the open sky are exposed to elevated carbon dioxide. Mean of 285 studies (Ainsworth and Rogers 2007).](http://ucanr.edu/sites/calagjournal/archive/?image=fig6302p68b.jpg)
Fig. 3. Net carbon dioxide assimilation (photosynthesis) as a function of carbon dioxide concentrations within a leaf (Ci) for C3 plants grown at either ambient (365 parts per million [ppm]) or elevated (567 ppm) atmospheric carbon dioxide concentrations, in free air CO2 enrichment (FACE) plots, where plants growing in soil under the open sky are exposed to elevated carbon dioxide. Mean of 285 studies (Ainsworth and Rogers 2007).

Fig. 4. Each line shows change in biomass over time for specific plants grown at elevated (567 ppm) and ambient (365 ppm) carbon dioxide atmospheres, in free air CO2 enrichment (FACE) plots (Dukes et al. 2005; Korner 2006) and open-top chambers (Rasse et al. 2005; Kimball et al. 2007).

Fig. 5. Differences in yield, aboveground biomass, leaf nitrogen (N) concentrations and grain protein concentrations between C3 plants grown at elevated (567 ppm) and ambient (366 ppm) carbon dioxide concentrations under heavy (high N) and normal N fertilization (low N). Symbols and error bars designate means ± 95% confidence interval for crops (Ainsworth and Long 2005), trees (Curtis and Wang 1998), grasses (Wand et al. 1999), all plant species (De Graaff et al. 2006) and grain protein (Taub et al. 2008). Parentheses contain number of studies included in the meta-analysis.
The balance between C3 carbon fixation and photorespiration depends on the relative amounts of carbon dioxide and oxygen entering the active site of rubisco (i.e., portion of the enzyme involved in the primary chemical reactions) and the affinity of the enzyme for each gas (i.e., degree to which it attracts carbon dioxide or oxygen). At current atmospheric levels of carbon dioxide and oxygen (about 380 and 209,700 ppm, respectively), photorespiration in most crops (C3 plants including wheat, rice, barley, oats, legumes, vegetables, and fruit and nut trees) dissipates) over a quarter of the organic carbon produced during carbon dioxide assimilation (conversion from inorganic to organic form) (Sharkey 1988).
In contrast, C4 crops (such as corn, sorghum and sugar cane), which have a metabolic carbon dioxide pump that increases the concentration of this compound at the catalytic site of rubisco, minimize photorespiration at the expense of the additional energy required for pumping.
Elevated levels of atmospheric carbon dioxide inhibit photorespiration in C3 plants, making photosynthesis more efficient. Initially, this accelerates both their photosynthetic carbon dioxide assimilation and their growth by about a third. After a few days or weeks, however, carbon dioxide assimilation and growth both slow down until they are accelerated in the long term by only about 12% and 8%, respectively (figs. 3 and 4). Moreover, leaf nitrogen and protein concentrations ultimately decrease more than 12% under carbon dioxide enrichment (fig. 5). Such a loss of nitrogen and protein significantly diminishes the value of this plant material as food for animals and humans. Together these trends are known as carbon dioxide acclimation.

Wheat was grown in a controlled environmental chamber at elevated carbon dioxide (700 ppm). Plants in the three containers on the left received ammonium (NH4+) as the sole nitrogen source, whereas those on the right received nitrate (NO4−). Plants grown at ambient carbon dioxide under ammonium and nitrate nutrition were indistinguishable (not shown).
CO2 acclimation hypotheses
Severl hypotheses have been put forward to explain carbon dioxide acclimation.
Carbohydrate sink limitation.
According to this hypothesis, plants under carbon dioxide enrichment initially assimilate more carbon dioxide into carbohydrates than they can incorporate into their growing tissues. In response, they diminish carbon dioxide assimilation by decreasing their rubisco levels (Long et al. 2004). This change in rubisco levels, however, is not necessarily selective; the decrease may instead just be part of the overall decline in protein and nitrogen concentrations (Ainsworth and Long 2005; Makino and Mae 1999).
Progressive nitrogen limitation.
Another hypothesis for carbon dioxide acclimation is that shoots accumulate carbohydrates faster than roots can absorb nitrogen from soils, making leaf nitrogen concentrations decrease (Hungate et al. 2003; Luo et al. 2004; Norby et al. 2001; Reich et al. 2006). As these leaves senesce and drop to the ground, (1) plant litter quality declines, (2) microbial immobilization of soil nitrogen increases because of the high carbon-to-nitrogen ratios in the litter, (3) soil nitrogen availability to plants further diminishes because more soil nitrogen is tied up in microorganisms, (4) plants become even more nitrogen limited, (5) plant protein levels decline and (6) plant processes including photosynthesis slow down (fig. 6). This hypothesis, however, has difficulty in explaining the variation in carbon dioxide acclimation among sites (Finzi et al. 2007) and among methods of carbon dioxide enrichment (Ainsworth and Long 2005).
![Differences in leaf carbon fixation capacity (photosynthesis [PS]) versus total nitrogen concentration (N) between C3 plants grown at elevated (567 ppm) and ambient (366 ppm) carbon dioxide concentrations. Each symbol designates the mean ratio for a species. Shown are the regression line (solid, slope = 0.815, r = 0.71) and 1:1 line (dotted). This data suggests that changes in photosynthesis from carbon dioxide enrichment derive from changes in plant nitrogen levels under carbon dioxide enrichment (Ellsworth et al. 2004).](http://ucanr.edu/sites/calagjournal/archive/?image=fig6302p69.jpg)
Fig. 6. Differences in leaf carbon fixation capacity (photosynthesis [PS]) versus total nitrogen concentration (N) between C3 plants grown at elevated (567 ppm) and ambient (366 ppm) carbon dioxide concentrations. Each symbol designates the mean ratio for a species. Shown are the regression line (solid, slope = 0.815, r = 0.71) and 1:1 line (dotted). This data suggests that changes in photosynthesis from carbon dioxide enrichment derive from changes in plant nitrogen levels under carbon dioxide enrichment (Ellsworth et al. 2004).
Role of photorespiration
We have discovered another explanation for carbon dioxide acclimation: in C3 plants, shoot assimilation of nitrate into organic nitrogen compounds depends on photorespiration, so any condition that inhibits photorespiration (elevated carbon dioxide or low oxygen concentrations) also inhibits shoot nitrate assimilation (figs. 7 and 8). Thus, at elevated carbon dioxide concentrations, C3 plants that rely on nitrate as a nitrogen source suffer severe deprivation of organic nitrogen compounds such as proteins. The resulting decline in organic nitrogen compounds reduces the plants' yield and biomass production. While high applications of nitrogen fertilizer may partially compensate for this, the plants' nitrogen and protein concentrations still diminish (fig. 5).
Ammonium and nitrate are the two main sources of nitrogen that are accessible to plants from the soil. Plants show a wide range of responses to carbon dioxide enrichment because the balance between nitrate and ammonium availability varies over seasons, years, locations and plant species. In an annual California grassland where nitrate was the predominant nitrogen source, net primary productivity diminished under carbon dioxide enrichment (fig. 4) (Dukes et al. 2005). This was presumably because elevated carbon dioxide inhibited plant nitrate assimilation (by both shoots and roots), and the grasses became deprived of organic nitrogen. In contrast, ammonium is the major form of nitrogen available to plants in marshes because wet, anaerobic soils promote denitrification (the conversion of nitrate into nitrous oxide and dinitrogen gas) and nitrate leaching (the removal of dissolved nitrate into deep groundwater or surface water). For example, the dominant C3 plant in the Chesapeake Bay marsh (Scirpus olneyi) showed little carbon dioxide acclimation (fig. 4); even after a decade of treatment, photosynthesis and growth remained about 35% greater under carbon dioxide enrichment (Rasse et al. 2005), with little change in nitrogen concentrations (Erickson et al. 2007). In wheat, another C3 plant, elevated carbon dioxide atmospheres stimulated less growth under nitrate than under ammonium nutrition (fig. 9; see photo, page 69).
Physiological mechanisms
Several physiological mechanisms appear to be responsible for the dependency of nitrate assimilation on photo-respiration.
First, the initial biochemical step of nitrate assimilation is the conversion of nitrate to nitrite in leaves. This step is powered by the high-energy compound NADH (reduced nicotinamide adenine dinudeotide), and photorespiration
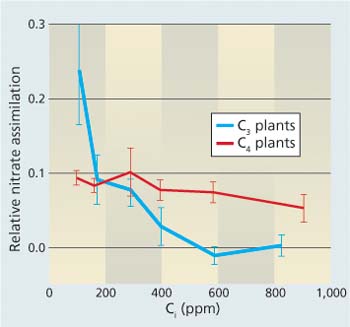
Fig. 7. Response of nitrate (NO3−) assimilation in C3 and C4 plants as a function of carbon dioxide concentrations inside a leaf (Ci). Relative NO3− assimilation was assessed from changes in CO2-O2 fluxes with a shift from NH4+ to NO3− nutrition (ΔAQ). The C3 species included barley (Bloom et al. 1989), wheat (Bloom et al. 2002), tomato (Searles and Bloom 2003), Arabidopsis (Rachmilevitch et al. 2004) and Flaveria pringlei and giant redwood (Bloom, unpublished data). The C4 species included maize (Cousins and Bloom 2003, 2004) and Flaveria bidentis and Amaranthus retroflexus (Bloom, unpublished data).
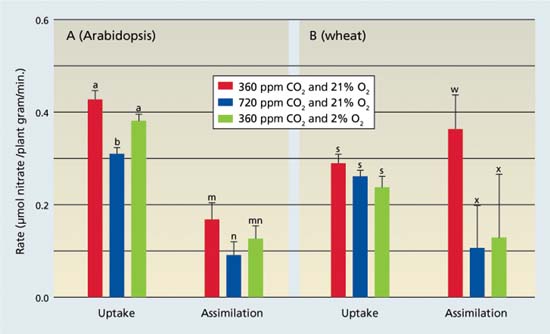
Fig. 8. Nitrate (NO3−) uptake as the amount of NO3 depleted from a medium, and nitrate assimilation as the difference between the rates of net NO3− uptake and net accumulation of free NO3− in plant tissues: (A) 36-day-old Arabidopsis or (B) 10-day-old wheat were exposed to 360 ppm carbon dioxide (CO2) and 21% O2, 720 ppm carbon dioxide and 21% O2, or 360 ppm CO2 and 2% O2. Shown are the mean ± SE (n = 13-16). Treatments labeled with different letters differ significantly (P < 0.05). Light levels were 500 and 1,000 micromoles of quanta per meter squared per second for Arabidopsis and wheat, respectively (Rachmilevitch et al. 2004).

Environmental chambers in the UC Davis Controlled Environment Facility are helping scientists to understand how plants react to changes in atmospheric carbon, oxygen and other greenhouse gases.

Twelve wheat seedlings were subject to atmospheres containing various concentrations of carbon dioxide and oxygen. The bubbles are a thin water layer that lines the top of the chamber to control its temperature.

Fig. 9. Biomass (grams dry mass) and leaf area (cm2) per plant of wheat seedlings grown for 14 days in controlled environment chambers at 360 or 700 ppm carbon dioxide and under NH4+ or NO3− nutrition. Shown are mean ± SE for four replicate experiments, each with 8 to 10 plants per treatment. Treatments labeled with different letters differ significantly (P< 0.05) (Bloom et al. 2002).
Second, the subsequent biochemical step of nitrate assimilation is the conversion of nitrite to ammonium in the cloroplasts of leaf cells, which requires the transport of nitrite into the chloroplast. Elevated carbon dioxide inhibits this transport (Bloom et al. 2002).
Third, this subsequent step also requires chemical energy from the oxidation of a different high-energy compound called ferredoxin. Several other processes — in particular, carbon dioxide assimilation — depend on the same energy source and seem to have priority in using it. Ferredoxin becomes involved in nitrate assimilation only when carbon dioxide availability limits C3 photosynthesis (Backhausen et al. 2000; Peirson and Elliott 1988).
Carbon dioxide and food quality
Many crops in California depend on nitrate as their primary nitrogen source. As atmospheric carbon dioxide concentrations rise and nitrate assimilation diminishes, these crops will be depleted of organic nitrogen, including protein, and food quality will suffer (Taub et al. 2008). Wheat, rice and potato provide 21%, 14% and 2%, respectively, of protein in the human diet (FAOSTAT 2007). At elevated carbon dioxide and standard fertilizer levels, wheat had 10% less grain protein (Fangmeier et al. 1999; Kimball et al. 2001). Similarly, grain protein in rice (Terao et al. 2005) and tuber nitrogen in potato (Fangmeier et al. 2002) declined by about 10% at elevated carbon dioxide concentrations.
Several approaches could mitigate these declines in food quality under carbon dioxide enrichment. Increased yields may compensate to some degree for total protein harvested (fig. 5). Several-fold increases in nitrogen fertilization could eliminate declines in food quality (Kimball et al. 2001), but such fertilization rates would not be economically or environmentally feasible given the anticipated higher fertilizer prices and stricter regulations on nitrate leaching and nitrous oxide emissions. Greater reliance on ammonium fertilizers and inhibitors of nitrification (microbial conversion of ammonium to nitrate) might counteract food quality decreases. Nevertheless, the widespread adoption of such practices would require sophisticated management to avoid ammonium toxicity, which occurs when plants absorb more of this compound than they can assimilate into amino acids and free ammonium then accumulates in their tissues (Epstein and Bloom 2005). Several of these issues might be simultaneously addressed by fertigation, or frequent additions of small amounts of ammonium-based fertilizers in water delivered through microirrigation.
These findings have broad implications for the future of plant distributions and food production. Enriched carbon dioxide atmospheres will not enhance the performance of C3 plants to the extent originally envisioned. A 10% decline in food protein content will further burden regions of the world already affected by hunger. With a better understanding of ammonium and nitrate use by crops and careful nitrogen management we can turn these phenomena to our advantage.
As carbon dioxide rises, food quality will decline without careful nitrogen management

