


All Issues
Monitoring shows vegetation change at multiple scales
Publication Information
California Agriculture 55(6):42-47. https://doi.org/10.3733/ca.v055n06p42
Published November 01, 2001
PDF | Citation | Permissions
Abstract
Several historical data sets from the UC Hopland Research and Extension Center (HREC) provide valuable information on vegetation dynamics at multiple spatial and temporal scales. An early botanical survey by Harold Heady and Al Murphy provides a baseline for examining landscape-level changes in species richness and distribution over 50 years. We conducted a floristic survey between 1995 and 1999 and found gains and losses of native and non-native species abundance across the field station. On two sites where sheep were removed in 1958, permanent transects provide valuable information about plant community responses to protection from livestock grazing; in the oak understory, native perennial blue wildrye increased steadily, while in grasslands native purple needlegrass was less abundant after 43 years. On a cleared watershed, originally diverse in hardwood species, we found that only interior live oak and coast live oak were significantly reestablished after 30 years.
Full text

Historic vegetation surveys at the UC Hopland Research and Extension Center were used to describe changes to the plant communities over several decades. Density of blue wildrye (Elymus glaucus) on this understory transect in Riley Ridge Biological Area increased from 0.17 plants/square meter in 1979 (top) to 6.54 plants/square meter in 2000 (bottom). Standing herbaceous biomass increased considerably in 2000.
Change is ubiquitous on hardwood rangelands, from dramatic alterations in climate (Bartolome 1989a), grazing fauna (Edwards 1996) and human impacts over the past few millennia. In the 19th century, waves of European and American immigration into California — and the concurrent introduction of livestock production, mining and plowing — led to widespread and rapid introduction of non-native, mostly European annual plant species (Heady 1977; Hamilton 1997). Not much is known about these introductions or what native rangeland species were lost because they were, for the most part, undocumented. A lack of baseline vegetation data has prevented scientists from monitoring the extreme changes that occurred, severely constraining retrospective interpretation. Monitoring of vegetation change during the second half of the 20th century, however, has been more successful.
Although planning and design are an important part of any monitoring program, historic data that were not necessarily intended for monitoring purposes can nonetheless be quite useful. For example, historical data sets are available from the UC Hopland Research and Extension Center (HREC) covering nearly 50 years. Data are available at several spatial scales ranging from large (landscape) to small (individual plants). These data sets, which include permanent photo stations, are only now beginning to yield their full potential. Shortly after UC purchased the HREC in 1951, station superintendent Al Murphy and UC Berkeley range management professor Harold Heady began an intensive floristic inventory, which has provided a baseline for current flora comparisons across the landscape.
In 1958, Heady established two sheep exclosures and placed permanent transects in them to monitor the density of perennial grasses. These transects allowed scientists to monitor small-scale vegetation change by resampling over 40 years. In addition, we examined patterns of large-scale change at a watershed level after its woody vegetation was cleared in the early 1960s. Together these studies show how vegetation has changed at widely varying scales of analysis.
The spatial and temporal dimensions used to measure changes in vegetation cover and composition greatly affect the patterns detected. Measurements taken at the finest grain of spatial resolution are usually restricted to small areas, while large-scale studies examine areas of greater extent at a coarser resolution. For example, small-scale studies using vegetation transects and grazing exclosure plots provide high levels of accuracy, but the resulting information cannot be generalized beyond the area under study. Examining historic records of species presence and absence over time for large geographic ranges such as HREC provides a general picture of change but sacrifices precision. Changes in vegetation at varying spatial scales can be detected over short time intervals such as between seasons or over longer periods spanning years. Examination of vegetation change at multiple spatial and temporal scales provides a more complete understanding of the topic. HREC's unique and intact hardwood rangeland and abundant natural habitats, combined with an infrastructure that allows for a variety of agricultural and ecological research, are valuable assets for scientists seeking insights from both shortand long-term vegetation monitoring studies.
Vegetation at HREC
Lying approximately 40 miles inland from the Pacific Ocean, the HREC experiences a typical Mediterranean climate of hot, dry summers and cool, wet winters. Because it encompasses nearly 3,000 feet of elevation change with numerous soil types and diverse topography, as many as 650 vascular plant species occur within a rich suite of vegetation types and plant communities (Murphy and Heady 1983). Mixed oak woodlands, hardwood forest and grasslands occupy most of the lower two-thirds of the center, while chaparral is more common at higher elevations. Nutrient-poor serpentine outcrops provide a refugia for unique native plant communities. Together these habitat types support a diverse assemblage of species.
Hardwood trees at HREC are among the most diverse in the state and include eight species of oak, along with California bay, madrone, big-leaf maple, California buckeye and white alder. Using the most recent classification system for California's hardwood rangelands (Allen et al. 1991), at least 12 hardwood cover types (subseries) have been identified at HREC. At the higher elevations, chaparral is more prevalent on south-facing slopes, whereas a dense, forested canopy of Douglas fir, California nutmeg and canyon live oak occur on steep north-and east-facing slopes. Scattered across the field station are numerous seasonal creeks, sag ponds, vernal pools and springs, all providing a diversity of wetland habitat. Recent inventories have added 51 moss, three liverwort and 70 lichen species to the nonvascular flora at HREC.
Large-scale trends
Murphy and Heady's initial botanical survey documented all the vascular plants known to occur within HREC boundaries, providing a reference collection for land managers, researchers and students. Between 1952 and 1954 nearly 450 species were recorded, and over the next 30 years the number of species grew to 600 (Murphy and Heady 1983). In addition, they estimated the abundance of many species collected in the early 1950s. In 1983, abundance estimates were reassigned, this time to the entire flora. Collectively, these data, which are available on herbarium specimen labels, allowed us to examine changes in species richness, abundance and distribution across the field station.
One study resurveyed Murphy and Heady's sites and documented species gains and losses as well as changes in abundance (Heise and Merenlender in press). Field surveys between 1995 and 1999 did not relocate 43 species in the original record, although our efforts added 53 new species to the flora. Much of the species loss we documented can be attributed to habitat conversion or displacement by invasive species. Over half of the species that could not be relocated occurred initially in chaparral or wetland habitat (fig. 1). This can be partially explained by range improvement experiments in the 1960s and 1970s that converted some of the chaparral collection sites to grassland (Murphy 1976, 1978). In addition, some of the wetland sites where collections were taken are row dominated by pennyroyal (Mcniba puJegium), an invasive non-native plant that was uncommon when it was first collected in 1969.
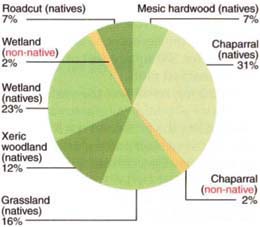
Fig. 1. Habitat types of plants that could not be relocated during 5-year survey between 1995 and 1999 at HREC.

Fig. 2. Habitat types of new plant species found during 5-year survey between 1995 and 1999 at HREC.

Watershed II at Hopland before it was cleared, left; in 1969, about 10 years after vegetation was cleared and burned, center; and in 2001, right. Thirty years after clearing, only one black oak and seven blue oaks were established, but extensive live oak and riparian vegetation is returning. Photos by Al Murphy and Colin Brooks.
Of the 53 new species found since 1995, about half were from wetland habitats (fig. 2). Of these, two-thirds were native. New species found in grassland and xeric (dry) woodland habitats also consisted of native and non-native species. In contrast, only native species were found in rock outcrops, mesic (moist) hardwood forest and chaparral. These results are similar to findings on other hardwood rangelands in California where non-native species favor more disturbed habitat (Knops et al. 1995).
A consequence of large-scale sampling is a loss of accuracy in estimating changes in abundance. In order to make valid comparisons in our later surveys, we adopted the coarse terms of relative magnitude, such as sparse, occasional and numerous, which were used in the early 1950s and 1980s. Using these terms, we detected only drastic changes across time. Three non-native grasses in particular, barb goatgrass (Aegilops triuncinlis), big quakinggrass (Briza maxima) and hedgehog dogtail (Cynosurus echinatus), which were uncommon or absent in the 1950s, became widespread and abundant by the mid 1990s. Barb goatgrass is especially troubling because of its ability to invade serpentine sites.
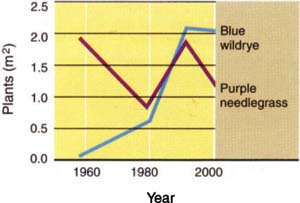
Fig. 3. Changes in native blue wildrye (Elymus glaucus) density on 10 permanent transects located under blue oak (Quercus douglasii) canopy and changes in native purple needlegrass (Nassella pulchra) density on 11 transects in open grassland. Sheep grazing was eliminated in 1958. Transects were sampled in 1959, 1979, 1991 and 2000.
Changes in perennial grass
In 1956, two areas of mixed grassland and oak woodland at the HREC were fenced to exclude sheep. The Riley Ridge Biological Area is about 250 acres in size at an elevation of 3,000 feet on the east boundary of the station. Foster Biological Area is smaller (62 acres) and at lower elevation (985 feet) on the southwest edge of the station. In 1958, Heady established four clusters of three permanently marked transects, 100 feet long and 5 feet wide, in each area where significant numbers of native perennial grasses existed; the clusters and transects were not randomly located. Changes over time should reflect the response of one component of vegetation to the removal of sheep at the plot-scale level (Bartolome 1989b) for two types of sites: open grassland and blue oak understory. All perennial grasses rooted in the transects were tallied in 1958, 1979, 1991 and 2000. This data set provides a picture of changes in the herbaceous layer over 43 years for 500 square feet of permanently marked plots.
A striking result was the variation in density of perennial grasses over time among the grassland transects (fig. 3). The overall pattern of change was a decline by 1979 and a return to original density by 1991. On several grassland plots, the non-native annuals medusahead (Taeniatherum caputmedusae) and Italian thistle (Carduus pycnocephalus) dominated in 1979 (Bartolome and Gemmill 1981). By 2000, perennial grass density was higher than in 1979, but still lower than in 1959 immediately after transect establishment and sheep exclusion, and before Italian thistle became common. Purple needlegrass (Nassella pulchra), which was previously suggested as a species adapted to disturbance and not protection from grazing, is the most common perennial (Bartolome and Gemmill 1981).
In contrast, native grasses in the oak understory transects increased strikingly between 1959 and 1991, particularly after 1979 (fig. 3). That increase stopped between 1991 and 2000. The oak understory is now dominated by blue wildrye (Elymus glaucus), which apparently benefited from the exclusion of grazing sheep after 1958.
This result illustrates the problems in application of the standard equilibrium model for vegetation change on hardwood rangelands and the value of long-term historical data sets. The generally accepted range-condition model does not accurately forecast the highly variable and unpredictable changes in perennial grass abundance in grasslands (Bartolome 1993). At Hopland, for example, the dominant species, purple needlegrass, was less abundant in 2000 than in 1958. After more than 40 years without livestock grazing, the density of native perennial grasses was lower in open grassland than when the transects were established, but considerably higher than in the late 1970s.
Since 1979, we have examined the transects for the presence of seedling oaks and infrequently found a few clusters of seedlings up to 2 inches tall, patchily distributed on the oak canopy transects. There have never been any larger seedlings on the transects and there is no evidence of successful oak recruitment. It is possible that changes in weather patterns have played an important role at these sites. We plan to investigate this theory.
Hardwood recruitment patterns
Much of the research on oak recruitment (the successful regeneration of seedlings and saplings) has focused on planting experiments designed to determine factors limiting seedling development (McCreary 2001), or on restoration projects involving the planting of thousands of oaks across many acres (Nives et al. 1991). Oak recruitment, or the lack thereof, has primarily been studied at the stand scale over short time spans, necessitating examination of seedling recruitment and development. Early studies of a cleared watershed at HREC offer the opportunity to monitor hardwood recruitment across a larger spatial and longer temporal scale. Data on overstory vegetation across a 200-acre watershed at HREC from 1958, 1965 and 1999 were used to examine the types and locations of hardwood establishment over 34 years.
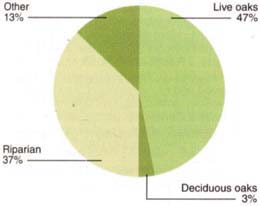
Fig. 4. Types of tree regeneration observed in HREC Watershed II since deforestation was completed in 1965.
Watershed II at HREC was the focus of a rangeland productivity and water-yield study that provided critical information on the original vegetation coverage, methods of vegetation removal and immediate results of the clearing (Heady and Pitt 1979). Typical of hardwood rangeland occurring in northwest California, the watershed has a drainage basin of 195 acres, ranges in elevation from 590 feet to 1,322 feet, and has a slope range of 2 to 39.6 degrees with a mean of 18.5 degrees. Soils are primarily of the Franciscan formation and approximately 3 feet thick. About 60% of the watershed is composed of south-, southwest- or southeast-facing slopes, and just under a quarter of the watershed is in north-, northwest- or northeast-facing slopes. The watershed has a long history of sheep grazing that continues today.
Most of the overstory vegetation in the watershed was cleared between 1959 and 1965 for the rangeland study. Heady and Pitt (1979) recorded vegetation characteristics before, during and immediately following the clearing and burning treatment. Historical photos account for the few hardwood trees that remained immediately after the type conversion and distinguish these from hardwoods that were established in the last 30 years. Recruitment of live oaks and riparian-dependent species was observed on the more mesic north- and east-facing slopes (fig. 4).
We found that 30 years after clearing only one black oak and seven blue oak trees were established. In contrast, extensive live oak and riparian vegetation cover is returning. Physiographic variables are strongly associated with the location of hardwood and shrub regeneration (Brooks and Merenlender 2001). The detection of these trends required an analysis across a large geographic area and historic records of the time and extent of vegetation clearing recorded by earlier researchers at HREC.
Across all scales
Monitoring vegetation change at multiple spatial and temporal scales allowed us to observe a variety of patterns. This is what we would expect since studies at the landscape scale reveal general patterns of vegetation change due to climate, topography and geology not captured in smaller areas. However, small plots monitored at HREC often include extensive amounts of environmental variation that can affect species turnover.
Monitoring of small (500-square-foot) plots revealed fine scale and unpredictable changes in native grass species abundance that were not mimicked in the abundance changes noted for the entire field station. For example, the understory perennial grasses, nearly all blue wildrye, showed a general and slow initial increase after 1959, rapid increase between 1979 and 1991, then slowed down over the past 9 years. Using the coarse measures of abundance inherent in the herbarium data set, no significant change was detected for blue wildrye and purple needlegrass after 50 years. This suggests that the observed changes in cover at the plot scale are site specific and may be related to the interaction of the site with removal of livestock, or with weather.
The increasing spread of non-native invasive species was detected on the smaller-scale open grassland plots and on the larger scale of the entire center. Some of these invasive species have come to dominate much of the open grasslands at HREC, even after the removal of grazing livestock. For example, Italian thistle increased in abundance on the permanent grassland plots as well as at various sites across the center, as noted from herbarium collections. This pattern contrasts with more mesic, forested sites where native grass cover increased in the ungrazed permanent plots, and new native species were detected throughout HREC.
Even this review of long-term vegetation change provides only a snapshot of the status of invasive, non-native species at HREC. In particular, the vegetation at HREC is certainly responding to recent weather conditions such as drought in the early 1990s and El Nino. Coordinated studies are needed across the state to examine and compare the dynamics of range grassland systems and the impact of environmental changes such as pollution and climate change.
Some annual grass species such as big quakinggrass have become wide spread across the entire region, whereas other species such as barb goatgrass are spreading at HREC at an accelerated rate compared to elsewhere ( see p. 47 ). We believe that the former invasion is due to near-term weather factors and the latter to sheep ranch management, in addition to differences in seed dispersal strategies. Such causal relationships can only be fully recovered through ecological monitoring across a large number of rangeland sites and owners.

Riley Ridge Biological Area (left of HREC property fence) showed dramatic increases in herbaceous biomass and shrub cover between 1952, left, and 1982, right. The native forb turkey mullein (Eremocarpus setigerus) is abundantly visible in the heavily used foreground grassland on HREC property (left of fence) in 1952, but no longer evident in 1982. The chamise (Adenostoma fasciculatum) shrubs in the foreground are heavily browsed in both photos. Photos by Harold Heady.
Few blue oak seedlings were found along the study transects and only seven blue oak trees were established across the entire area of Watershed II since the forest was cut 30 years ago. On the other hand, we observed significant live oak recruitment. As deciduous oaks die without a new cohort being established, active restoration may be required to revegetate blue oak habitat. The heterogeneity between stands, even for individual species such as blue oak (Swiecki et al. 1997), due to history, age structure and environmental variables, makes it difficult to identify a clear regeneration pattern through time or across space.
We have taken advantage of long-term monitoring studies at different spatial scales to show that both the herbaceous layer and the hardwood forests have changed over the past 50 years and are likely to continue to do so. To answer many of the questions raised by this and similar studies, it is essential to support research centers such as HREC, coordinate a network of research sites including the UC Natural Reserve System, and establish new long-term ecological research sites for California's hardwood range-lands. In addition, the preservation and maintenance of historic data sets (such as herbarium records, permanent plots and photos) provide essential information for ecological monitoring, a critical component of adaptive management.
Monitoring shows vegetation change at multiple scales

