


All Issues
Supporting evidence varies for rangeland management practices that seek to improve soil properties and forage production in California
Publication Information
California Agriculture 74(2):101-111. https://doi.org/10.3733/ca.2020a0015
Published online June 09, 2020
PDF | Citation | Permissions
Abstract
California is increasingly investing in policies and programs that promote soil stewardship on natural and working lands as a way to help achieve multiple goals, including improved forage production and climate change mitigation. To inform the growing expectations for rangeland management activities to promote such services, we conducted an evidence synthesis assessing how four commonly suggested practices (silvopasture, prescribed grazing, compost application and riparian restoration) affect a suite of soil properties and plant-related metrics throughout the state. We extracted data on soil properties that are potentially responsive to management and relevant to soil health. We also extracted data on aboveground forage production, forage nitrogen content and herbaceous species richness. Our search resulted in 399 individual soil observations and 64 individual plant observations. We found that the presence of oaks had the largest effects on soil properties, with soil organic carbon, microbial biomass and other measures of soil fertility increasing beneath oak canopies. The presence of grazing increased compaction and total nitrogen, and decreased pH. Compost applications did not significantly affect any of the measured soil properties, but did boost forage production. Due to a lack of published data, we were unable to characterize the influence of rangeland riparian restoration on any of the soil or plant metrics in our review.
Full text
Rangelands make up approximately 57 million acres of California's land area, with Mediterranean-type grasslands, shrublands and woodlands accounting for 30% of this estimate (FRAP 2018). The predominant economic use of California's rangelands is forage production, which supports a $2.6 billion cattle industry (CDFA 2018). These landscapes are also globally recognized as hot spots of biological diversity (Myers et al. 2000), and they provide an array of ecosystem services beyond food production, including water and nutrient cycling (Byrd et al. 2015), pollination (Chaplin-Kramer et al. 2011), carbon storage for climate change mitigation (Dass et al. 2018), and recreational opportunities (Plieninger et al. 2012).
As the land area of California's rangelands continues to shrink due to dramatic population growth and concomitant cropland and urban expansion (Cameron et al. 2014), societal demands from each acre are increasing. Private ranchers and public rangeland managers now desire, or are expected, to predictably optimize production of the full array of values and services (Boyd and Svejcar 2009; Ferranto et al. 2014; Plieninger et al. 2012) in a way that promotes ecosystem resilience and adaptation to global and regional pressures (Hruska et al. 2017; Sayre et al. 2012).

Over 3 miles of riparian area at Tolay Lake Regional Park, Petaluma, California, that have been actively restored by Point Blue Conservation Science's Students and Teachers Restoring a Watershed (STRAW) program. The white squares are plants installed by volunteers, including coast live oak, buckeye, coffeeberry, California rose and more.
Soils are receiving increased attention within the scientific discourse on rangeland management because of their role in supporting ecosystem services like forage production and, in some cases, climate change mitigation (Byrnes et al. 2018; Derner et al. 2016; Derner et al. 2018). In California, as elsewhere, this recognition is making its way into the public sphere, influencing both policy and practice (Bradford et al. 2019; Byrnes et al. 2017).
Indeed, a 2011 survey of California ranchers identified that maintaining or improving soil “health” was, on average, a midpriority goal that ranked similarly to managing weeds and water quality (Roche et al. 2015). Although the same survey revealed that managing soils to promote carbon sequestration was not an important goal for ranchers (Roche et al. 2015), since 2011 there have been several new policies and incentive programs put in place to promote carbon sequestration on rangelands. For example, rangeland soil management is now named by the California 2030 Natural and Working Lands Climate Change Implementation Plan as a critical climate change mitigation strategy, and policy and funding through the California Department of Food and Agriculture's Healthy Soils Program aim to maximize this outcome.
The growing expectation for range management activities to promote on-site (e.g., forage production) and public (e.g., carbon sequestration) ecosystem services demands a quantitative synthesis of the literature summarizing whether, and to what degree, desired outcomes have been shown to be achievable by implementing these activities. To that end, we synthesized the documented effects of four commonly prescribed rangeland management practices on a suite of soil properties and plant-related metrics in California. These management practices, which are currently incentivized by the Healthy Soils Program, are silvopasture establishment (USDA NRCS 2016, 381), prescribed grazing (USDA NRCS 2016, 528), compost application, and riparian restoration (USDA NRCS 2016, 391).
Previous work indicates that these practices have the potential to influence plant dynamics and increase soil organic carbon (hereafter soil C) in some rangeland ecosystems (Byrnes et al. 2018; Dybala, Matzek, et al. 2019; Gravuer et al. 2019) — and there is reason to believe they can also affect other soil properties, such as soil compaction and nitrogen (N) availability (Byrnes et al. 2018; Dahlgren et al. 1997; Zhou et al. 2017). In California's arid and semiarid Mediterranean climate, however, the effects of rangeland management may be overwhelmed by strong interannual variability in precipitation (Jackson and Bartolome 2007), spatial variability in soil type and topography (Booker et al. 2013; Graham and O’Geen 2016) and land-use history (Huntsinger et al. 2007).
This variability results in mixed evidence for California rangelands to respond to management practices and generates risks associated with extrapolating results from single studies or from other, perhaps more malleable, mesic regions (Allen-Diaz and Jackson 2005). Our targeted search and synthesis of California's literature at least partially addresses these risks, and supplements ongoing work in the state that is providing scientific underpinnings for an expanding rangeland management framework.
Literature review and synthesis
We performed a subject-wide evidence synthesis on data derived exclusively from California rangelands. A subject-wide evidence synthesis is a way to review and summarize the effects of multiple practices or interventions simultaneously using some combination of qualitative and quantitative methods, and it can be a cost-effective approach to support management and policy decision making (Shackelford et al. 2019). In our synthesis, we extracted soil and plant data from relevant studies that fulfilled specific requirements and aggregated similar response metrics into umbrella categories (e.g., the cations category combines values of individual exchangeable cations, base saturation and cation exchange capacity).
For composite metric-management pairs that had soil observations across at least three sites, we assessed the overall influence of management practices on each metric using random effects meta-analysis modeling, with log response ratios as the effect size (Hedges et al. 1999). For composite plant metrics, we took the same approach but relaxed the criteria to two (rather than three) sites so we could include the effects of compost amendments. Where possible, we also broke apart the composite categories to look at the effect of management practices on contributing variables (on individual exchangeable cations, base saturation, etc.) and explored whether soil texture and aridity moderated the response of metrics to management across studies.
Because we were interested in documenting published evidence in addition to identifying gaps for metric-management pairs regardless of their potential to be included in a formal meta-analysis, we retained all management practices regardless of their support in the literature and qualitatively described our findings in those cases where evidence was limited. See the technical appendix online for more details.
Levels of evidence: Highly variable across management practices
Our literature review resulted in 399 individual soil observations compiled from 37 publications, reports or unpublished data sets, and 64 individual plant community observations from 26 publications (table 1; table 2). The resulting data set represented 35 unique study areas located across California's Mediterranean-type grassland, shrubland and woodland systems (fig. 1); eight of these areas were UC reserves or other research stations, from which 71% and 50% of the soil and plant-related data sets were derived, respectively. Oak presence had the greatest number of soil observations (59% of the compiled data set), followed by grazing (33%), compost additions (5%) and riparian restoration (3%). This pattern held for plant metrics as well, with 44%, 34%, 13% and 9% of the observations associated with oak presence, grazing, compost additions and riparian restoration, respectively.
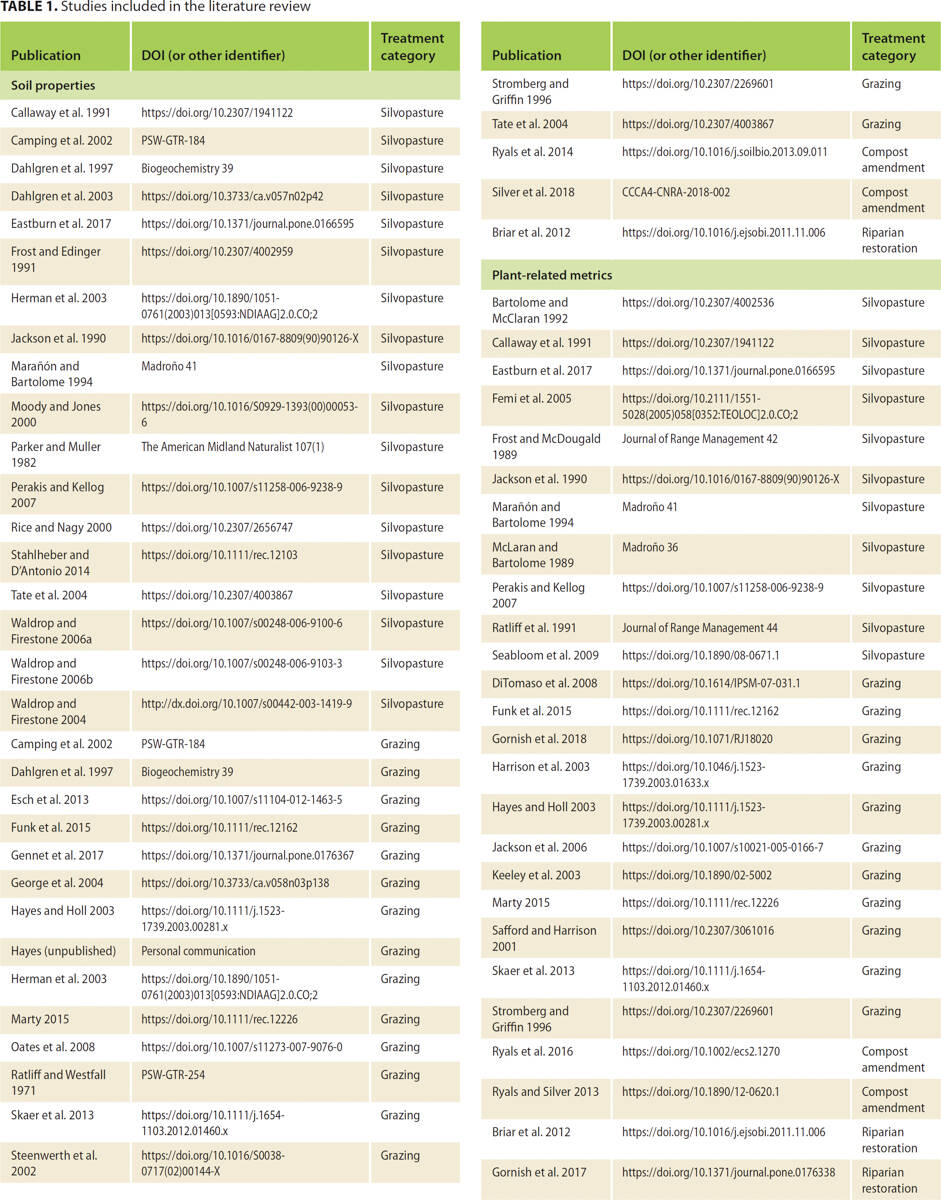
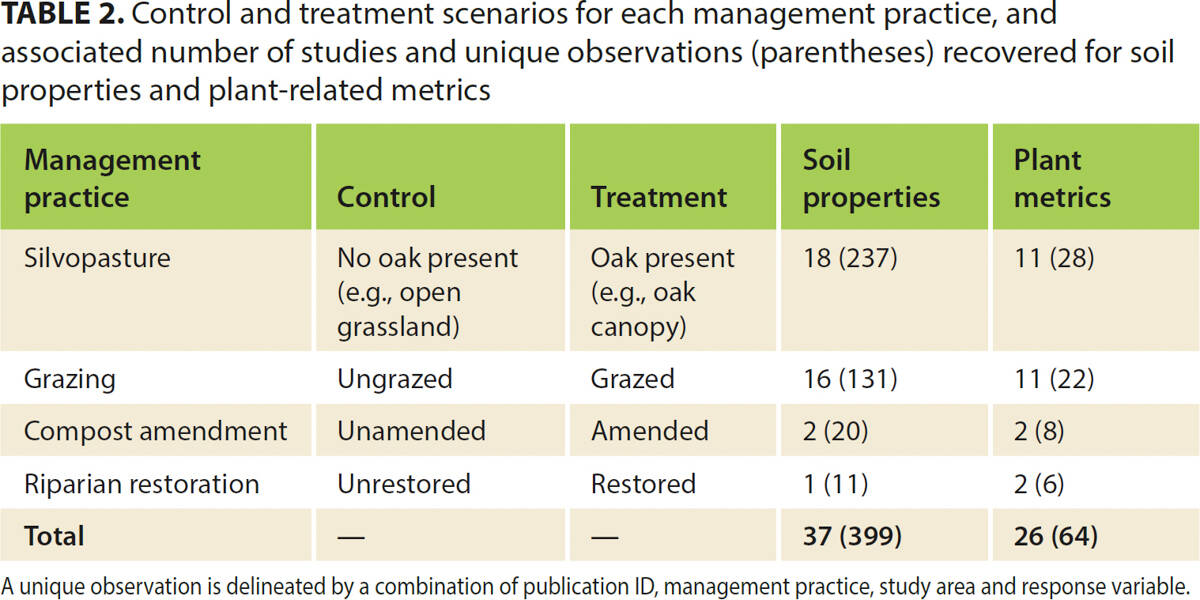
TABLE 2. Control and treatment scenarios for each management practice, and associated number of studies and unique observations (parentheses) recovered for soil properties and plant-related metrics
FIG. 1. Map of study sites included in the literature review, showing where soil properties and plant-related metrics were assessed.
Across all studies, the number of unique observations per metric varied from one to 93 (fig. 2), with soil N being the most frequently measured parameter, followed by cation availability and soil C content. Only 12 studies presented both soil and plant-related metrics, limiting our ability to draw direct relationships between the response of soil properties and plant dynamics to management intervention.
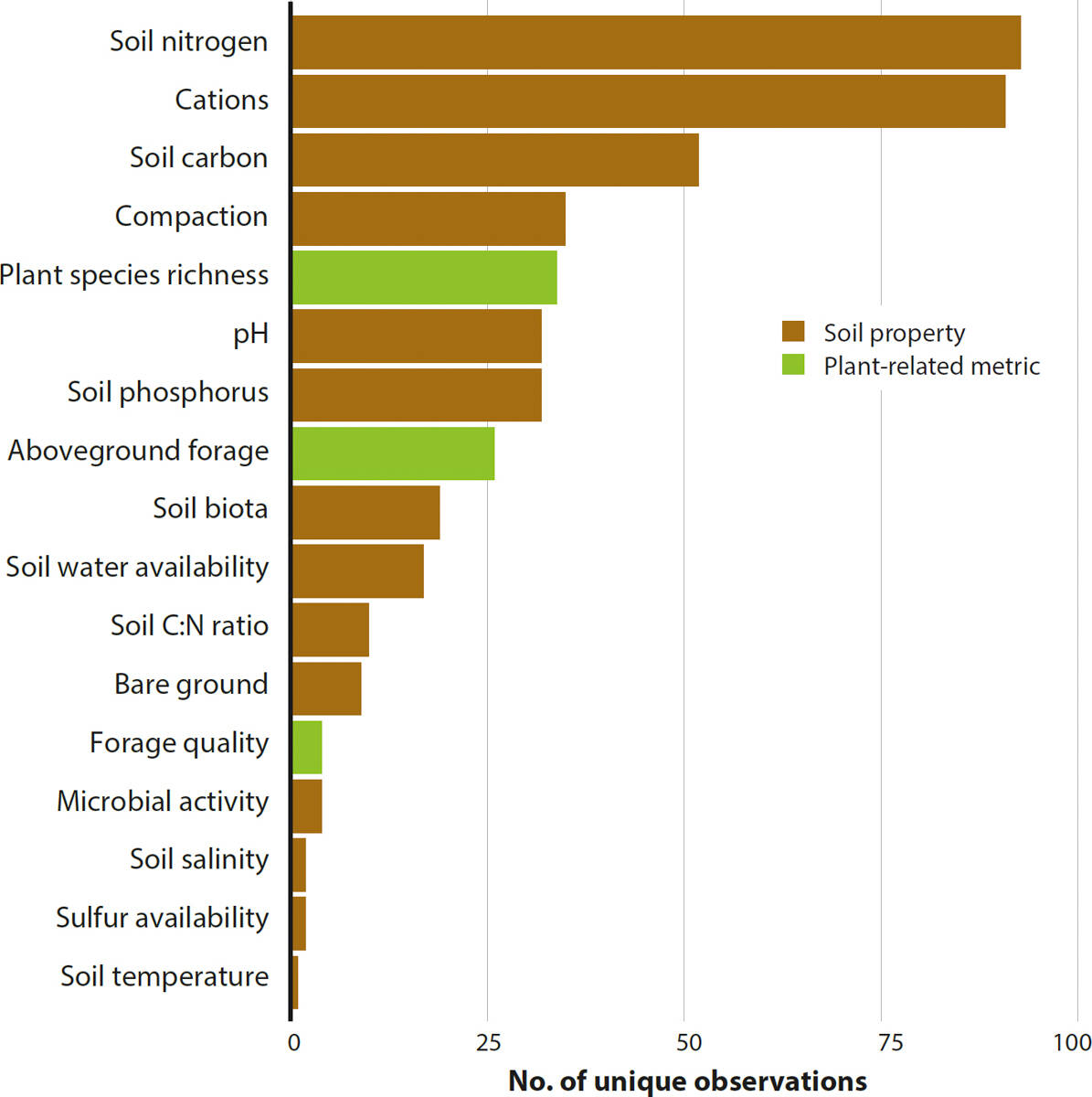
FIG. 2. The number of unique observations representing each soil property and plant-related metric recovered from the literature.
Silvopasture: Oaks enhance soil fertility
Silvopasture is an ancient practice that has been implemented in many regions around the world and is recognized for its potential to optimize economic and ecological production while building resilience into some working landscapes (Jose and Dollinger 2019). The Natural Resources Conservation Service defines silvopasture as the establishment or management of desired trees and forages with the purpose of providing improved forage production, shelter for livestock, soil quality and carbon sequestration, biological diversity and reduced erosion (USDA NRCS 2016).
In California's published literature, silvopasture establishment on rangelands is best represented by work on oak trees (Quercus spp.). And while oaks were historically clear-cut with the intention of maximizing forage production in California (a foundational assumption that was later challenged) (Huntsinger and Fortmann 1990), hardwood rangeland landowners increasingly value and manage for oaks, and the state is striving to increase the pace and scale of oak stewardship by tripling funding for oak savanna reforestation by 2030 (CNWL CCIP 2019; Huntsinger et al. 2010). As such, we focused specifically on assessing the effects of oak presence on soil properties and plant-related metrics in our review, and we included oak removal studies in addition to observational studies measuring conditions under oaks versus in nearby open grasslands.
We found the presence of oaks had the largest effects on soil properties of any of the management practices assessed. When the data were pooled across all studies and contexts, soils beneath oak canopies were more fertile (higher levels of soil phosphorus [P], soil N and cation availability), had greater amounts of soil C and microbial biomass, were less compacted and contained more soil moisture than nearby open grasslands (fig. 3). The effects of oaks on soil N were reflected in both the total N and plant-available N pools, and the effects on cations were largely driven by increases in exchangeable potassium, magnesium and calcium (fig. 4).

FIG. 3. Mean response ratio (InRR), number of unique observations and significance of management–soil property relationships. Significant difference between treatment and control for a given soil property is denoted by color, with blue points significant and red points insignificant. The size of each circle is a function of the number of papers supporting that relationship.
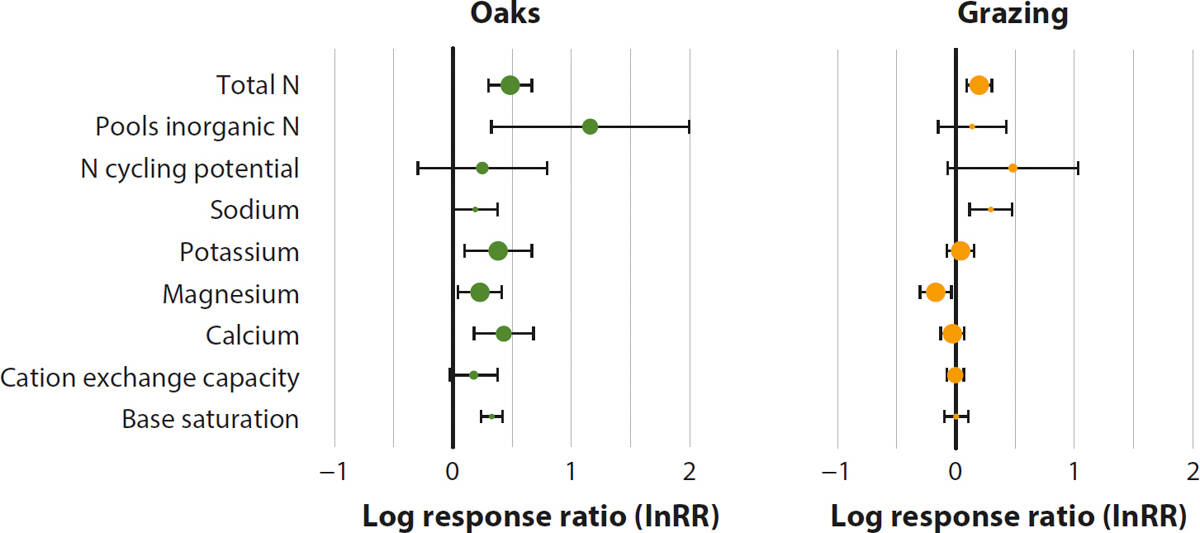
FIG. 4. Mean response ratio (InRR) and 95% confidence intervals (CI) for the response of soil N availability and individual cation metrics to oaks and the presence of grazing. If a metric has InRR > 0 and a 95% CI that does not overlap zero, it significantly increased in the presence of oaks or grazing. The size of each circle is a function of the number of unique observations supporting that relationship.
When accounting for environmental context, we found that soil texture moderated the effect of oaks on soil C such that the increase beneath oaks was amplified in soils with higher clay content (fig. 5). The effect of oaks on soil P was also greatest in finer-textured soils, and in less arid sites (data not shown). This context dependency aligns with expectations from theory and practice (Booker et al. 2013; Byrnes et al. 2018), suggesting that silvopasture-induced gains in soil C and some metrics of soil fertility, while still apparent, may be muted in sandy soils and hotter, drier areas.
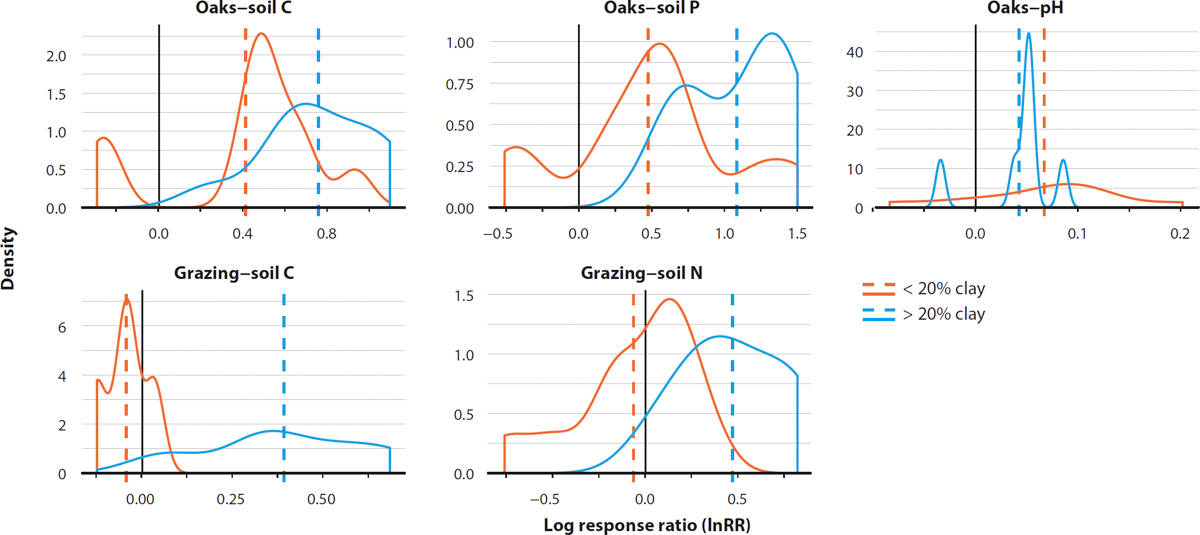
FIG. 5. Density plot displaying significant relationships between oak and grazing response ratios (InRR) and soil texture (percentage clay). Dashed lines = mean response ratio for a given texture category. Nine other management-soil property relationships were assessed and returned nonsignificant results (data not shown). The modulating effects of soil texture were also assessed for oaks on forage, and grazing on plant species richness. Neither relationship was significant (data not shown). Where possible, a similar exercise was performed to assess how aridity moderates management impacts. Aridity was found to only significantly moderate the effects of oaks on soil phosphorus levels and forage production (data not shown).
While it is possible that oaks preferentially establish on sites that are inherently more fertile, rapid declines in soil fertility have been observed following the removal of oaks, indicating that the trees create rather than respond to these conditions (Dahlgren et al. 2003; Herman et al. 2003). This “island of fertility” effect associated with oaks has been attributed to multiple potentially interacting mechanisms. These include the interception of wet and dry nutrient deposition by oak canopies (Callaway and Nadkarni 1991; Perakis and Kellogg 2007), greater litter inputs beneath oak trees (Dahlgren and Singer 1991; Knops et al. 1996), increased soil faunal activity (Dahlgren et al. 2003), hydraulic lift (Ishikawa and Bledsoe 2000) and increased root interception of nutrients that would otherwise be leached from the system (Perakis and Kellogg 2007).
It is also possible that oak roots relocate nutrients from adjacent open grasslands, concentrating them beneath their canopy — or that livestock elevate soil nutrients as they seek shade beneath these trees. However, both of these mechanisms appear unlikely (Dahlgren et al. 1997; Perakis and Kellogg 2007). Whatever the driving cause, it is clear that oaks create patches of soil fertility associated with increased levels of soil C and microbial biomass across California's rangelands, with possible consequences for public benefits such as climate change mitigation.
The effects of oaks on soil fertility did not seem to translate into similar effects on forage productivity and herbaceous species richness, at least with enough consistency to be significant when summarized across all studies and contexts (fig. 6). In the case of forage production, this is perhaps not surprising since considerable variability exists among individual oak trees, with some trees promoting forage production and others inhibiting it (Callaway et al. 1991).
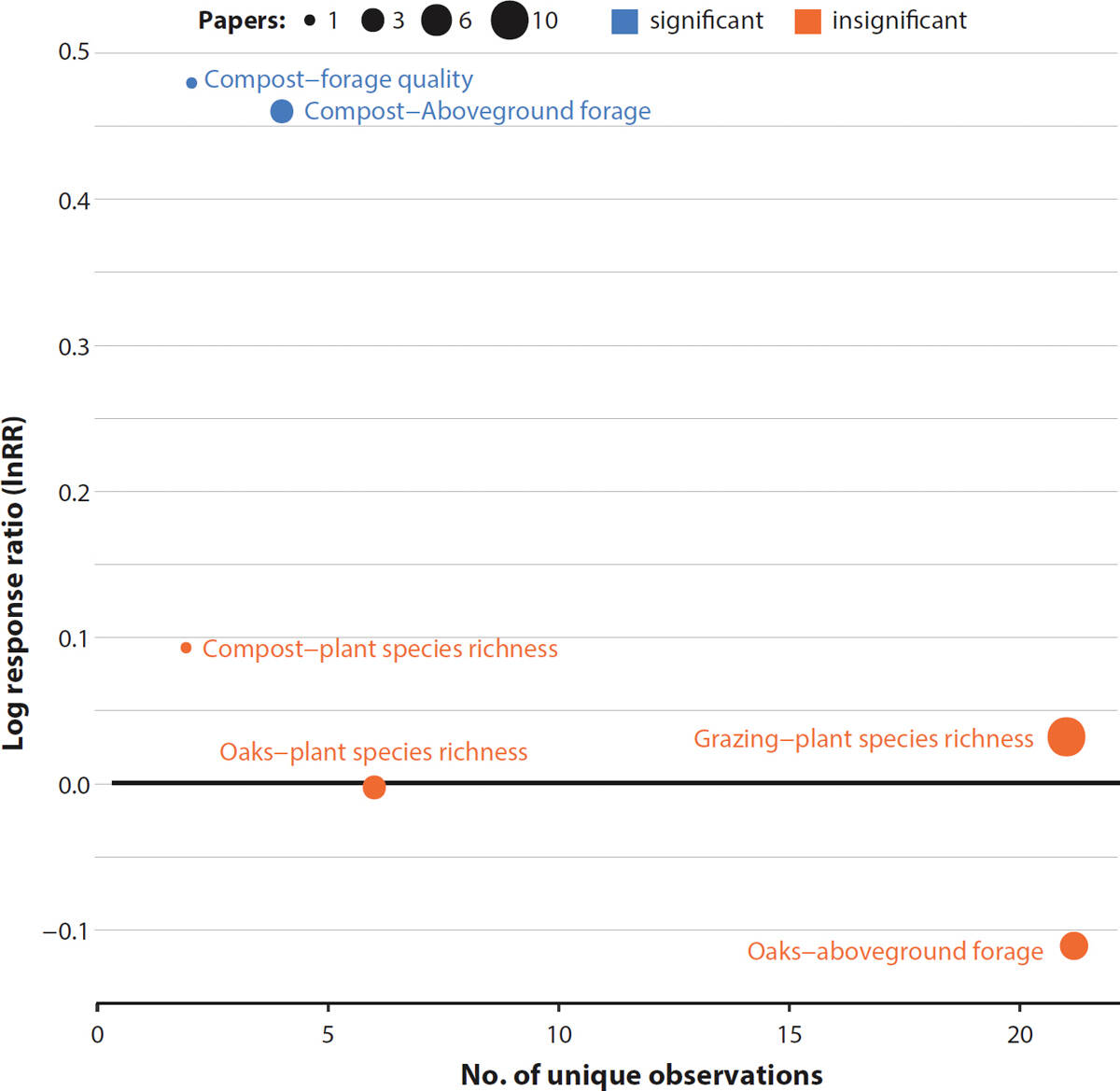
FIG. 6. Mean response ratio (InRR), number of unique observations and significance of management-plant metric relationships. Significant difference between treatment and control for a given soil property is denoted by color, with blue points significant and red points insignificant. The size of each circle is a function of the number of papers supporting that relationship.
Prior work has found oak individuals that inhibit understory production typically have shallower root systems, a phenomenon that may be due to any number of environmental or genetic reasons and which results in competition for limiting resources or suppression of herbaceous plant growth by allelopathic chemicals (Callaway et al. 1991; Koteen et al. 2015). At a watershed-scale, forage productivity may also show an inverse relationship with the density of oak trees (Battles et al. 2008). Despite the potential importance of these factors in driving oak-forage productivity relationships across the state, we were unable to account for either in our review. However, we were able to detect a relationship with aridity, with forage production decreasing beneath oak trees in cooler, wetter sites (data not shown).
A number of areas for future work stand out as priorities for this management practice. First, research is needed to quantify rates of change in soil properties with oak establishment, as the literature to date generally focuses on comparing established trees (> 80 years old) to adjacent open grasslands. Understanding how quickly and how much each soil property changes after an oak planting will be important for policy and practice, which have expectations on much shorter time scales (< 5 years).
At the same time, given the strong and relatively consistent influence of mature oaks on soil properties, priority should be placed on determining how to improve oak regeneration and ensure the success of silvopasture projects presently and in California's future climate (Bernhardt and Swiecki 2015). Finally, because forage dynamics are critical to livestock production systems, future work should continue disentangling patterns in forage productivity and bolstering evidence for oak impacts on forage quality.
Grazing: Soil texture affects response of soil C
In California's rangelands — which typically receive minimal inputs like irrigation and fertilizer — livestock grazing is the dominant land use and is therefore one of the most readily available management tools for landowners (Huntsinger et al. 2010). The ability of grazing to influence ecosystem outcomes particularly in water-limited systems is an area of active debate (Booker et al. 2013; Sanderson et al. 2020; Stanton et al. 2018; Teague et al. 2013). However, grazing management has been shown through meta-analysis to alter soil properties such as soil C, total N and bulk density globally (Abdalla et al. 2018; Byrnes et al. 2018) and plant community dynamics regionally (Stahlheber and D’Antonio 2013).
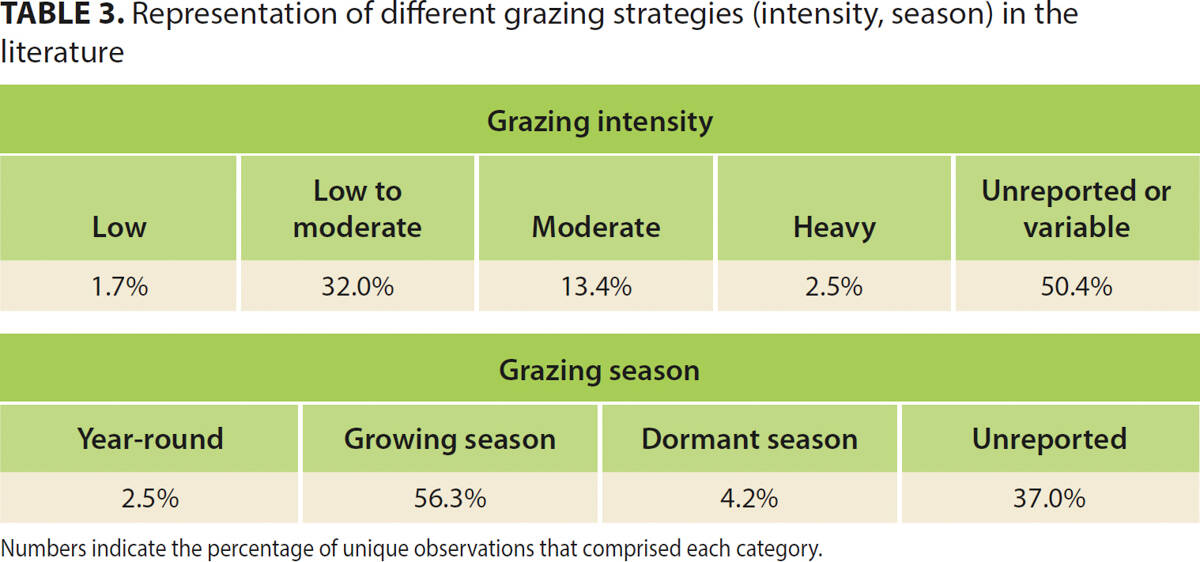
Unfortunately, although our intent was to assess the effects of different grazing strategies, we were unable to go beyond presence-absence of grazing. Over half of the grazing studies (63%) reported grazing intensity, and most of these reported grazing during the growing season at low-moderate or moderate intensities.
Observations of extreme grazing intensities (low and heavy) were lacking, as were studies documenting the effects of year-round and dormant-season grazing (table 3). The near absence of these strategies in the literature probably reflects the fact that very little land is managed this way in California (Huntsinger et al. 2007), although management will certainly vary from site to site based on resource needs and management goals, and some regions may rely on year-round grazing more than others (Liffmann et al. 2000).
The relative uniformity of grazing strategies across presence-absence studies, along with the lack of studies that explicitly manipulate and compare deferment and within-season rest and rotation (i.e., compare among “grazing systems”), prevented us from quantitatively assessing the effects of these finer-level, but potentially important, moderators.
Our literature review revealed that the presence of grazing, when pooled across all studies and contexts, significantly influenced a number of soil properties (fig. 3). Specifically, grazed sites had greater soil compaction and considerably more bare ground than ungrazed sites, a pattern that has been shown elsewhere (Augustine et al. 2012; Byrnes et al. 2018), with possible consequences for outcomes such as water infiltration (reduced infiltration) (Savadogo et al. 2007) and grassland breeding bird habitat (increased suitability) (Gennet et al. 2017).
Grazed sites also had higher levels of soil N, driven by a small but significant increase in total N (fig. 4), and lower levels of exchangeable magnesium (fig. 4) and soil pH (fig. 3). This collectively indicates that grazing can influence soil fertility across California's landscapes. Increases in soil N with grazing have been reported in some global reviews (Abdalla et al. 2018) but not others (Byrnes et al. 2018), and without more information we are unable to determine the cause of such an increase in California. However, possible explanations include changes in plant species composition, biomass production and allocation, and spatial redistribution of N with livestock waste (Piñeiro et al. 2010).
Policymakers are increasingly interested in leveraging grazing management to help sequester carbon and achieve climate change mitigation goals (Byrnes et al. 2017). While evidence is mixed and estimates are highly uncertain, prescribed grazing has been suggested to sequester carbon at rates up to 1.8 metric tons per hectare per year in some mesic regions (Conant and Paustian 2002). Whether the sequestered carbon remains in the ground long enough to help mitigate climate change depends on various aspects of the system, including how protected the carbon is from microbial decomposition (Lavallee et al. 2020).
In California, the influence of grazing on soil C is thought to be small relative to factors such as climate and soil type, with the net effect depending on a number of site-specific conditions such as management history and soil texture (Stanton et al. 2018). Supporting this expectation, and in line with prior work (Silver et al. 2010), we found that across all studies the presence of grazing had no significant effect on soil C.
However, similar to oaks, we found that soil texture significantly modified the effect of grazing, such that grazing had more of a positive effect on soil C in soils with higher clay content (fig. 5). McSherry and Ritchie (2013) report a similar pattern for arid sites in their global meta-analysis. Possibly, sandier soils are more susceptible to C loss through reductions in vegetation cover and increased wind erosion of fine particles (Steffens et al. 2008). Finer-textured soils also have a greater capacity to stabilize increased inputs of organic material that may occur through compensatory growth of vegetation (Jackson and Bartolome 2007; Singh et al. 2018).
While this result is derived from a relatively small sample size and should therefore be interpreted with caution, it suggests that soils with higher clay content, such as those of the Central Coast and Sacramento Valley, may be more amenable to grazing-induced improvements in soil C — and other measures of soil fertility (fig. 5) — than those with lower clay content, such as soils of the San Joaquin Valley. Future work should focus on understanding whether, and to what degree, this relationship holds true with an expanded data set across the state.
A number of priorities for future grazing research emerged from our review. Specifically, robust long-term and well-replicated experiments are needed to assess how soil properties and plant metrics co-vary in response to grazing strategies across California's diverse rangeland landscapes. These experiments should aim to go beyond presence-absence and begin identifying the impact of strategies such as within-season rest and rotation. Soil properties that may deserve elevated attention include microbial biomass and soil moisture (or other proxies of soil water availability), as these lacked enough evidence to be included in the current review and yet are important components of semi-arid Mediterranean-type systems.
Finally, while some work has been done to assess the effects of residual dry matter — a proxy for grazing intensity — on forage productivity (Bartolome et al. 2007), few studies directly compare the effects of grazing strategies on this plant metric; therefore, measurements of forage productivity and quality in response to different grazing strategies are needed.
Compost amendments: Applications increase forage
Applying composted organic amendments to rangeland soils is a practice increasingly promoted by government and nongovernment agencies across the state. These amendments have been shown to improve on-ranch soil C sequestration and forage quantity and quality at two sites in Northern California (Ryals et al. 2014; Ryals et al. 2016). In addition, a recent global meta-analysis by Gravuer et al. (2019) showed that rangeland forage production and soil C increase in the first few years following application of organic amendments. However, Gravuer et al. (2019) caution that these benefits may come with trade-offs depending on the type of organic material used and the rates of application.

An experimental plot at TomKat Ranch, Pescadero, California, receives a onetime compost application as part of a larger statewide NRCS project to evaluate the effects of compost on soil- and plant-related outcomes.
If the potential on-site ecosystem benefits of compost amendments lead to increased demand for compost across the state, that could drive greater diversion of organic wastes away from traditional fates such as landfills and help to mitigate greenhouse gas emissions downstream as well (DeLonge et al. 2013). For these reasons, expert opinion ranks rangeland compost amendments as one of the most promising ways to help achieve statewide climate change mitigation goals (Stanton et al. 2018), and this practice is currently incentivized through the Healthy Soils Program. The expectation by many, although perhaps not all (Booker et al. 2013), is that these amendments will improve forage dynamics for livestock production systems while simultaneously promoting climate adaptation and resiliency of the state's working landscapes (Flint et al. 2018).
In our review, we were able to assess the effects of compost on soil C and total soil N, and from two sites we were able to assess plant-related metrics. We found that forage production and quality increased with compost addition (fig. 6) but that soil C and total soil N remained unchanged (fig. 3). The lack of significant differences in these soil properties possibly reflects the limited amount of observations derived from within California and also the short duration in which effects have been tracked over time. For example, one of the primary soil C data sets contributing to our review came from samples collected just 1 year after compost was applied (Silver et al. 2018).
Given more time, changes in soil properties may start to emerge (Gravuer et al. 2019). Indeed, modeling efforts have demonstrated the potential for compost to improve on-site and public ecosystem benefits across California over longer time frames (Flint et al. 2018; Ryals et al. 2015; Silver et al. 2018). However, well-replicated long-term studies are needed to validate those models, improve the evidence base for compost amendments to achieve desired outcomes across California's diverse rangelands, and determine whether the currently limited supply of compost would provide greater benefits in other systems, such as irrigated cropland.
Riparian restoration: Limited published evidence
Riparian corridors are globally recognized for their potential to provide multiple values and services, including carbon sequestration (Dybala, Matzek, et al. 2019), wildlife habitat (Dybala, Steger, et al. 2019; RHJV 2004) and maintenance of water quality and quantity (Bedard-Haughn et al. 2004; George et al. 2011). In California, where more than 90% of riparian ecosystems have been lost or degraded (RHJV 2004), state investment in riparian restoration is set to triple by 2030 (CNWL CCIP 2019).
Expectations from both private and public sectors are that riparian restoration will help to recoup lost ecosystem services and build resilience and adaptation into California's landscapes (Jackson et al. 2015; Seavy et al. 2009). However, evidence documenting the benefits of riparian restoration on California rangeland soils and understory vegetation is lacking in the literature (Jackson et al. 2015; Matzek et al. 2018). As a result, we were unable to characterize the influence of rangeland riparian restoration on any of the soil properties or plant metrics in our review.
The few studies that exist present mixed evidence for this practice's influence on ecosystem properties on California rangelands. For example, Lewis et al. (2015) demonstrated considerable soil C accrual following a combination of active (e.g., planting) and passive (e.g., grazing removal) restoration of riparian forests in Marin, Napa and Sonoma counties. That pattern has also been recently demonstrated with riparian restoration of agricultural lands in the Central Valley (Dybala, Steger, et al. 2019). Moreover, in rangelands of Marin, Sonoma and Mendocino counties, Gornish et al. (2017) found higher native herbaceous plant richness in actively restored sites than in control sites. In contrast, Briar et al. (2012) found that 3 years of passive restoration had limited effects on soil nematode communities in Yolo County.
Adopting monitoring protocols and performing research to supplement implementation will be critical to help bolster peer-reviewed evidence, constrain estimates on the rate and magnitude of change over time and provide the groundwork for evaluating the effects of this rangeland management practice on soil properties and plant metrics across California.

Left, fifth grade students learn how to plant acorns as part of a riparian restoration project in Sonoma County, California. Right, coast live oak acorns are protected from herbivory after planting in Sonoma County.
Three takeaways
Three general takeaways emerge from our quantitative synthesis of California's literature: (1) rangeland management signals can be observed for some soil and plant-related metrics but not others, suggesting that climate and inherent spatial variability in soil type, topography and land-use history can, but do not always, overwhelm the effects of management on California's rangelands; (2) soil texture moderates how some soil properties respond to management, and so it may be an important contextual variable to consider in modeling activities and land management decisions; (3) levels of in-state evidence that support our understanding of these four management practices vary by metric, but overall fall in this descending order: oaks > grazing > compost amendments > riparian restoration.
Continued support for research on these management practices will help to inform recommendations and adoption at scale. Predicting the effects of each management approach could further be supported by the refinement of organizing frameworks such as ecological site descriptions and state-and-transition models (Jackson and Bartolome 2002; Ratcliff et al. 2018), which are currently less developed for California than other Western states. In addition to more traditional research approaches, demonstration projects supported through the Healthy Soils Program offer one promising avenue to gain additional data, as does monitoring associated with state-funded projects through the California 2030 Natural and Working Lands Climate Change Implementation Plan.
Supporting evidence varies for rangeland management practices that seek to improve soil properties and forage production in California

