


All Issues
Invisible invaders: Insect-transmitted viruses threaten agriculture
Publication Information
California Agriculture 52(2):23-28. https://doi.org/10.3733/ca.v052n02p23
Published March 01, 1998
PDF | Citation | Permissions
Abstract
The vast movement of people and agricultural products between distant geographical regions has created unprecedented opportunities for introducing plant viruses and the insects that carry them (vectors) to new areas. Outbreaks of new viruses may be favored in these agroecosystems by crop susceptibility, the presence of particular weeds and certain agricultural practices. In some cases, conditions in these ecosystems may be ideal for the emergence of altered plant viruses and new virus/vector relationships. This may result in the appearance of insect-transmitted plant viruses in crops and regions where they have not been seen before. Because plant viruses and their insect vectors are intimately linked, the status of both must be considered in formulating strategies to prevent or slow their introduction, as well as to manage any invasions. To illustrate these points we highlight two situations that could threaten California agriculture. First, a devastating plant virus, tomato yellow leaf curl geminivirus, is not present in California, but an insect (the silverleaf whitefly) that transmits it is present. Second, the brown citrus aphid is not present in California, but a citrus virus (citrus tristeza closterovirus) that this insect efficiently spreads, is present.
Full text
Despite technological advances leading to tremendous yield increases for many crops, modern agricultural production continues to face pest threats, among them insects, plant pathogens, and weeds. Often growers are faced with multiple pests, which exacerbates crop damage and complicates management strategies. A dramatic example of how two types of pests “team up” to cause major problems for California agricultural production is the case of plant viruses and their insect vectors.
Plant viruses are foreign genetic elements that take over a plant's cellular machinery and use it to produce their own genetic material and proteins. This has dire consequences for the plant, altering many normal plant functions such as cell division and photosynthesis, thus resulting in disease. While symptoms of virus infection vary depending on the particular virus and plant, they generally involve stunted and distorted growth, changes in leaf coloration and shape, and poor flower and/or fruit yield and quality. Most plant viruses are not stable outside of living plant cells and so need a way to spread from infected plants to uninfected plants. Plant-feeding insects are ideal agents for spreading plant viruses because of their high rates of reproduction, dispersal abilities, and obligate use of particular plants as food. Indeed, intricate relationships have coevolved among insects, viruses and the plant hosts they share, resulting in remarkably efficient spread of viruses from plant to plant.

Symptoms of tomato yellow leaf curl geminivirus in leaves of an infected tomato plant. Below, plants infected at a young age are severely stunted and new shoots grow straight up, resulting in small, compact plants with bushy tops that are commonly referred to as ‘broccoli’ or ‘bonsai’ plants.
California farmers are already faced with a plethora of insect-transmitted viruses that can limit crop production, and for which there are few effective management options. While the viral diseases California growers currently face can be severe, they pale in comparison to some that have emerged in other areas of the world.
When a virus, either “new” or previously existing, causes sudden epidemics in a region, it is often called an emerging virus. There are various possible reasons underlying the emergence of diseases caused by plant viruses: ecological conditions may have favored the evolution of new viruses or new vector-virus relationships (such as the recent appearance of tomato mottle geminivirus in Florida associated with the introduction of silverleaf whitefly, Bemisia argentifolii); there may have been an expansion of the geographic range of existing plant viruses, insect vectors and susceptible crop plant species; or the emerging viruses may have been either unknown or relatively unimportant until changing agricultural practices or altered vector biology (for example, changes in host preferences or population dynamics) favored their dramatic increase. The unprecedented movement of people, agricultural products and propagules (such as seeds, bulbs, tubers and transplants) also provides tremendous opportunities for plant viruses and the insects that spread them (vectors) to move between widely separated geographic regions. Clearly, an awareness of these threats and the actions needed to avoid introducing these exotic pests is critical to all involved in California agriculture.
The completion of the proposed contained research facility on the UC Davis campus would provide academic, government and private industry researchers the opportunity to conduct important research on these exotic pests that could lead to new management strategies.
In this article, we will review the current status of two diseases caused by insect-transmitted plant viruses that have wreaked havoc in many parts of the world, and could someday pose a threat to California crop production. In the first case, tomato yellow leaf curl, the virus is not present in California, but the vector insect, silverleaf whitefly, is well established. In the second case, the opposite is true: citrus tristeza closterovirus is present in California but an efficient vector insect (the brown citrus aphid, Toxoptera citricida) is not.
Tomato yellow leaf curl virus
Tomato yellow leaf curl geminivirus (TYLCV) may be the most damaging virus to tomatoes because plants infected at an early stage of growth fail to produce fruit. While TYLCV-infected tomato plants produce abundant flowers, the virus causes flowers to fall off (abscise) long before setting fruit. In the field, yield losses can reach 100%. TYLCV gets its name from the fact that infected leaves are yellow except for the veins, show strong upward curling of the outer leaf margins and are small and crumpled. Plants infected at a young age are severely stunted and new shoots grow straight up, resulting in small, compact plants with bushy tops that are commonly referred to as ‘broccoli’ or ‘bonsai’ plants. In general, the younger the plant at time of infection, the more severe the stunting. The relative age at which a plant was infected can be determined because only the new growth will show symptoms.
TYLCV is a geminivirus (Geminiviridae family), a group of plant viruses characterized by twinned icosahedral virus particles, which look like two identical-sized soccer balls stuck together and from which the family name is derived (the Latin word gemini means twin). Gemini viruses can be carried by either leafhoppers or whiteflies, and whitefly-transmitted geminiviruses are one of the major emerging groups of plant viruses worldwide. For instance, whiteflies spread various tomato-infecting geminiviruses in Central and South America, Asia, India and the Middle East. While TYLCV and other whitefly-transmitted geminiviruses that infect tomato are not present in California, a whitefly that transmits these viruses is present. (The vector is silverleaf whitefly. See related article, p. 29 ).
In contrast to many diseases caused by whitefly-transmitted gemini-viruses, tomato yellow leaf curl is not a new problem. This disease was first described in Israel around 1940, and was associated with outbreaks of the sweetpotato whitefly (Cohen and Antignus 1994). Tomato yellow leaf curl was subsequently described in parts of the Middle East, Africa, Southern Europe, India and Asia. The causal agent of tomato yellow leaf curl was identified as a geminivirus in 1988 and it is now known that a number of distinct geminiviruses cause tomato yellow leaf curl-like symptoms in different parts of the world.
Although TYLCV is not present in California, it has recently become established in the Caribbean. By the late 1980s, the Dominican Republic had a flourishing processing tomato industry. Enough tomatoes were grown to supply the local canneries that produced tomato paste for the country's nearly 7 million people. However, in the fall of 1992, unusual virus-like symptoms were noticed on tomato plants in the northern production areas, and during the 1992–93 growing season entire seed beds and fields were lost. One year later, in 1993, these viral symptoms were observed in southern production areas as well. By the 1993–94 growing season, losses had reached millions of dollars, and the tomato acreage in the Dominican Republic had fallen by more than 75%. Canneries stayed in production by importing tomato paste but small farmers who depended on tomato production for much of their living were devastated and the tomato production areas became economically depressed.
Based on the association of the problem with high populations of whiteflies and the severity of the symptoms, the disease was suspected to be caused by a whitefly-transmitted geminivirus. The predominant disease symptoms were stunted growth and leaf yellowing and upcurling, which differed greatly from the symptoms associated with known Western Hemisphere tomato-infecting geminiviruses such as tomato mottle geminivirus in Florida and tomato leaf crumple geminivirus in Mexico.
In January 1994, a team of geminivirus researchers, R.L. Gilbertson (UC Davis) and D.P. Maxwell (University of Wisconsin-Madison), visited the Dominican Republic and noticed that the symptoms resembled those caused by TYLCV. Using molecular genetic techniques, the researchers confirmed that the mysterious tomato disease in the Dominican Republic was indeed caused by TYLCV. By 1997, the researchers also showed that the DNA sequence (the genetic code) of the Dominican Republic virus was 98% identical to that of an Eastern Mediterranean isolate of TYLCV (Nakhla et al. 1994; Polston et al. 1994). This virus had previously been reported only in the Eastern Hemisphere and had probably been inadvertently introduced into the Dominican Republic through infected plants or through whiteflies carrying the virus on plant material (such as tomato cuttings or transplants).

The silverleaf whitefly is a vector of tomato yellow leaf curl in other countries.
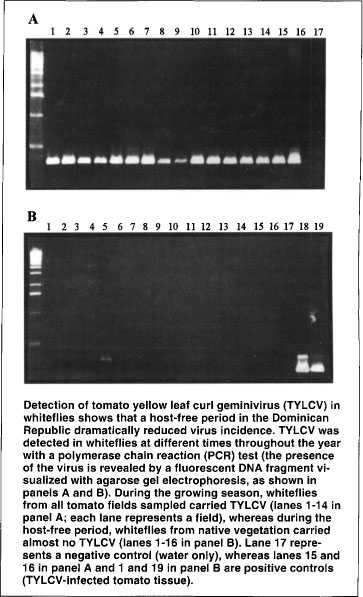
Detection of tomato yellow leaf curl geminivirus (TYLCV) in whiteflies shows that a host-free period in the Dominican Republic dramatically reduced virus incidence. TYLCV was detected in whiteflies at different times throughout the year with a polymerase chain reaction (PCR) test (the presence of the virus is revealed by a fluorescent DNA fragment visualized with agarose gel electrophoresis, as shown in panels A and B). During the growing season, whiteflies from all tomato fields sampled carried TYLCV (lanes 1–14 in panel A; each lane represents a field), whereas during the host-free period, whiteflies from native vegetation carried almost no TYLCV (lanes 1–16 in panel B). Lane 17 represents a negative control (water only), whereas lanes 15 and 16 in panel A and 1 and 19 in panel B are positive controls (TYLCV-infected tomato tissue).
Once the identity and distribution of the virus infecting the tomatoes was known, the Dominican Republic Ministry of Agriculture and the processing tomato industry formulated a strategy to revive the once-flourishing industry. The strategy's key component was prohibiting whitefly host crops (such as tomatoes, beans, melons and peppers) from being grown between the end of June (which is when the processing tomato season ends) and the end of September. Whitefly populations and TYLCV were further reduced by plowing under old tomato fields. During this host-free period, whitefly populations and the incidence of TYLCV in whiteflies decreased dramatically.

Citrus tristeza virus is not a serious problem in San Joaquin Valley citrus, but could become so if the brown citrus aphid is introduced.

Symptoms of CTV can be severe as shown here, where a ‘Valencia’ scion on a sour orange rootstock has been Infected.
Other successful components of the strategy included planting TYLCV-resistant tomato varieties and hybrids and using insecticides (primarily the systemic insecticide imidacloprid) to manage whitefly populations on tomatoes. As a result, the Dominican Republic's processing tomato industry has rebounded dramatically, and the 1996–97 growing season provided record yields that kept canneries from importing tomato paste for the first time in 6 years.
However, it is unlikely that TYLCV will ever be eradicated from the Dominican Republic. Moreover, TYLCV has spread from the Dominican Republic, possibly via long-distance movement of viruliferous whiteflies (those carrying the virus), to neighboring Caribbean countries including Haiti, Jamaica and Cuba. The virus was also found for the first time in Florida in 1997, where it was initially detected in tomato transplants sold in retail stores for home gardens (as seedlings in pony-packs). This raises the disturbing possibility that TYLCV has been inadvertently spread throughout the state via infected tomato transplants. However, whether the virus will become established in Florida remains to be seen.
TYLCV is clearly a potential threat to California's tomato industries, particularly because one of its vectors (the silverleaf whitefly) is well established in Southern California and seems to be expanding northward. Thus, introducing TYLCV into an area with tomato production and high whitefly numbers could result in the rapid spread and establishment of the virus in California. Once established, TYLCV would probably be difficult if not impossible to eradicate from California due to the diversity of crops grown and the wide plant host range of the silverleaf whitefly. These factors would make it difficult to implement solutions that have helped elsewhere, such as the host-free period in the Dominican Republic.
There are at least three possible ways new whitefly-transmitted geminiviruses could enter/appear in California: (1) introduction via infected plant material or viruliferous whiteflies on plant material; (2) introduction via flights of viruliferous whiteflies from areas with whitefly-transmitted geminiviruses; and (3) evolution of new geminiviruses from native forms. However, the most likely way TYLCV could enter California is via infected plant material. Thus, measures should be taken to prevent movement of plant material, especially tomato transplants, into California from areas known to have TYLCV or other whitefly-transmitted geminiviruses.
It is also important to be able to rapidly detect and characterize TYLCV and other whitefly-transmitted geminiviruses. We have developed a number of DNA-based rapid detection tests for TYLCV and other geminiviruses. Anyone seeing tomatoes with TYLCV-like symptoms should send samples to R.L. Gilbertson at the Department of Plant Pathology, UC Davis, for testing; results can be obtained in 24 to 48 hours.
If the virus did become established in California, it would be imperative to develop TYLCV-resistant tomato varieties. This is being approached using conventional breeding as well as biotechnology methods.
Citrus tristeza virus
The most economically important virus disease of citrus in the world is tristeza, which means sadness in Portuguese. Tristeza is caused by citrus tristeza virus (CTV), which belongs to the Closteroviridae, a family of plant viruses that have extremely large genomes (amounts of genetic material). Their large genomes are thought to contribute to the tremendous variability of CTV strains. Plant responses to CTV vary widely: some have such mild symptoms that they are not readily discernible while others undergo quick decline and death. CTV may have a severe impact on citrus when susceptible rootstock/scion combinations are infected. Because the severity of the symptoms depends so greatly upon the citrus cultivar and the rootstock/scion combination (Garnsey et al. 1987), the relationship between symptom severity and CTV strain diversity remains unclear. The virus can be spread by grafting of infected budwood, but CTV is most commonly dispersed by aphids. Several potential vectors, including Aphis gossypii, Toxoptera aurantii and Aphis spireacola colonize citrus in California. Aphis gossypii has been implicated as the principal vector of CTV in California (Dickson 1957), but is thought to be relatively inefficient in spreading the virus (Dickson et al. 1951, 1956; Grafton-Cardwell, Polek and Ullman, unpublished). Variation in CTV may alter how aphids transmit the virus, but the relationship is not fully known (Raccah et al. 1978).
CTV has been present in Southern California since 1939, killing more than 3 million trees. Incidence of CTV remains high in Southern California and many areas are under quarantine, prohibiting movement of any citrus planting material beyond quarantine borders.
CTV was first detected in the San Joaquin Valley (SJV) in the early 1960s but most citrus trees there have mild or nondiscernible symptoms, even though 15% to 20% of them are on the highly CTV-susceptible sour orange rootstock (Dodd and Gumpf 1987). The reason for this is still unknown. One possibility is that CTV strains transmitted by aphids in the SJV are mild and remain so over time. A more likely possibility is that the low incidence of severe tristeza in the SJV is largely due to the continuous removal of CTV-infected trees there by the Central California Tristeza Eradication Agency, as well as regulatory efforts preventing movement of citrus planting and propagative materials from areas under quarantine, and use of virus-free nursery budwood from the Citrus Clonal Protection Program.
CCPP: preventive medicine for citrus
Most California citrus groves planted in the last 40 years have an important connection to the University of California. They were established using budwood from the virus-free citrus collection developed and maintained by the Citrus Clonal Protection Program (CCPP).
The program, headquartered at UC Riverside, includes a UCR quarantine facility and a foundation block of trees located at the Lindcove Research and Extension Center.

All commercial citrus trees consist of a scion bud grafted on a selected rootstock. Rootstocks are chosen for their suitability to the soil in question, their disease resistance or other characteristics; the scion budwood is the actual fruit variety (navel orange, grapefruit, lemon etc.).
As is true in other commodities, citrus growers and researchers frequently import new commercial varieties from other citrus-growing areas, or exotic varieties for use in breeding programs to obtain trees with more desirable production characteristics. Since most of the important and destructive diseases of citrus are budtransmitted, it is imperative that imported budwood be tested, and that any exotic disease be eliminated, before release into the United States. An example would be the elimination of severe stains of the tristeza virus from budwood, as discussed in the accompanying article.
The CCPP was begun when the state's citrus growers, UC, and the California Department of Food and Agriculture entered into partnership in 1953. It has been continuously funded by the Citrus Research Board. Today the protected CCPP foundation block at the UC Lindcove Research and Extension Center has more than 700 trees representing 200 varieties of citrus.
The CCPP was the first program of its kind in the citrus industry. At present, it is also the only such program in the United States with a USDA Animal and Plant Health Inspection Service permit and the expertise to import, test and release clean citrus budwood. Most of the procedures and protocols used by the CCPP have been developed by program scientists, and they are the accepted standards for citrus quarantine, testing and treatment in the United States. They have also served as a model for the development of similar programs worldwide.
D. Gumpf is Plant Pathologist and heads the CCPP at UC Riverside.
The SJV is one of the few citrus-growing regions in the world that can grow citrus without intensively battling losses to CTV. However, this could be changed by the introduction of the brown citrus aphid (Toxoptera citricida), a CTV vector. The introduction of the brown citrus aphid into California would be problematic because: (1) this aphid is the most efficient vector of CTV, transmitting the virus at a rate 6 to 25 times greater than A. gossypii (California's current principal CTV vector); (2) this aphid has been associated with rapid spread of CTV strains causing severe symptoms in many citrus cultivars; and (3) the honeydew this aphid excretes during feeding causes serious damage to citrus due to sooty mold formation (reviewed in Yokomi et al. 1994).
The brown citrus aphid, native to China, is widely distributed throughout the citrus-growing regions of Asia, India, Australia, New Zealand, the Pacific Islands including Hawaii, sub-Saharan Africa and South America. The introduction of the brown citrus aphid to South America was blamed for the rapid spread of CTV isolates that killed tens of millions of trees during the epidemics of the 1930s and 1940s (reviewed in Yokomi et al. 1994). In recent years, the brown citrus aphid has also been introduced into Central America and the Caribbean, where it has spread rapidly. In 1995, it was detected in Florida and is now found on citrus throughout the state.
Given the rapid geographic expansion of the brown citrus aphid, the presence of this insect in Central America and Florida brings it dangerously close to the borders of California. While the brown citrus aphid has not been detected in California crops, it has been intercepted at ports of entry by the California Department of Food and Agriculture (CDFA).
There are several current regulatory and educational efforts aimed at preventing introduction of the brown citrus aphid and new CTV strains, to prevent spread of severe CTV strains within California, and to eradicate CTV in those regions where virus incidence is relatively low. For instance, the CDFA recently increased surveillance for the brown citrus aphid on agricultural products entering the state. To help ensure the success of this effort, citrus personnel have been trained to identify the brown citrus aphid in educational workshops conducted by Drs. Raymond Yokomi (USDA/ARS Fresno) and Elizabeth Grafton-Cardwell (Cooperative Extension entomologist, UC Riverside). Another effort to limit CTV in California is the Citrus Clonal Protection Program, which supplies virus-free nursery budwood and is under the leadership of David Gumpf, UC Riverside plant pathologist. This program has been critical to reducing spread of CTV isolates through grafting.
Current methods for managing CTV in areas where the brown citrus aphid is established include using chemicals and biological control agents to suppress the aphids, and using mild CTV strains to cross-protect citrus trees against infection by severe strains. However, both approaches have limitations. Efforts to suppress the brown citrus aphid may be inadequate because even very small numbers of aphids carrying the virus can cause severe epidemics. The drawback of cross-protection is that it works only against closely related isolates and/or strains, and CTV is so genetically diverse that cross-protection could fail to protect against the many unrelated severe strains. Also, severity is a relative measure of damage, and under some conditions the cross-protecting strains may be more severe than is acceptable to growers.
One hope for combating CTV and the brown citrus aphid may lie in introducing genes into citrus trees that will make them resistant to these pests. For example, introducing the gene for the CTV coat protein could protect citrus trees against infection by the virus. Other viral genes that might protect citrus trees include those that would interfere with the normal replication of the virus, or produce compounds that interfere with the aphid's transmission or acquisition of CTV to and from plants. Another possible strategy would be to identify genes that give citrus trees resistance to the brown citrus aphid, by preventing feeding and establishment, for instance. These genes could be transferred into citrus by traditional breeding or through biotechnology.
Invisible invaders: Insect-transmitted viruses threaten agriculture

