


All Issues
Evidence for a new species of whitefly: UCR findings and implications
Publication Information
California Agriculture 47(1):7-8.
Published January 01, 1993
PDF | Citation | Permissions
Abstract
The whitefly that caused over $500 million in damage to U.S. agricultural production in 1991 initially was referred to as a strain of sweetpotato whitefly. New studies provide evidence that this whitefly is a distinct species, now referred to as silverleaf whitefly. The research leading to this finding and the recognition of this new species are critical to California agriculture and directly impact the search for management strategies against silverleaf whitefly.
Full text
Producers of many food and fiber crops grown throughout the southern United States faced a common enemy in 1991 in the form of high densities of an introduced whitefly. The impact on such a wide diversity of crop plants (including alfalfa, broccoli, cabbage, cantaloupe, cauliflower, cotton, cucumber, tomato, squash, peanut, pepper, potato and watermelon) over such a large geographical area (Arizona, California, Florida, Georgia, Louisiana, New Mexico, and Texas) by one arthropod pest species in a single year was unprecedented. Agricultural losses in 1991 have been estimated at over a half billion dollars. In addition, close to 10,000 jobs, comprised mainly of migrant harvesting crews whose livelihood depends on seasonal agricultural production, were lost. Consumers in certain areas of the U.S. experienced shortages and price increases of fall and winter produce.
Based on morphological similarity to the sweetpotato whitefly, Bemisia tabaci (Gennadius), the severely damaging whitefly became known as the “B strain” of that species. The sweetpotato whitefly “A strain” has been present in the U.S. for many years without causing losses of the magnitude experienced in 1991. This fact, combined with other differences between the strains (California Agriculture, November-December 1991), has caused scientists to question whether the two whiteflies are strains of the same species or different species.
We conducted a number of experiments to determine how closely related the two whitefly strains were (Perring et al. 1993. Science 259: 74–77). Crossing experiments and mating behavior studies were performed to determine if the two strains were reproductively isolated. We also conducted genomic studies to evaluate the degree of phenotypic and genotypic similarity of the two whiteflies. In the crossing experiments, no female offspring were produced whenever A strain was crossed with B strain in any combination of parental sexes. Both male and female offspring were produced when males and females from like strains were crossed. Whiteflies exhibit haplo-diploidy, that is, males are produced from unfertilized eggs and females are produced from fertilized eggs. Consequently, this experiment indicated that, for inter-strain crosses, either no fertilization of eggs occurred or all female progeny died before maturing to the adult stage.
Upon discovering the lack of female offspring in crosses of strain A and strain B whiteflies, studies of whitefly mating behavior were initiated to determine the mechanisms of this apparent mating incompatibility. Male and female whiteflies of the same strain completed courtship behavior and copulated. Although whiteflies from different strains exhibited courtship behavior, none of the pairs copulated. Therefore transfer of gametes between strain A and strain B whiteflies was inhibited at the behavioral level.

Our genomic studies utilized two techniques: isoelectric focusing to evaluate differences in enzyme structure (see sidebar) and DNA differentiation studies using polymerase chain reaction amplification (PCR). Results from both of these studies indicated specific, fixed genomic differences between the whitefly types. These results support the biological data that indicated that these morphologically similar whiteflies represent different species.
Discussion
The results of the crossing, behavioral, and genomic studies provide evidence that the two whiteflies are distinct species; these results are consistent with previously recognized biological differences between the A and B strains. Because the newly recognized species causes a disease in squash known as squash silverleaf (California Agriculture, November-December 1991), we have recommended that it be referred to as silverleaf whitefly. Other common names for the silverleaf whitefly that have been used previously include the Florida colony, Florida strain, poinsettia strain, poinsettia whitefly, B type, and B strain.
Our research findings are pertinent, not only in the development of pest management strategies for the silverleaf whitefly, but in the broader context of national and global agriculture. On a global level, there is a need for a worldwide systematic revision of the genus Bemisia. Our studies utilize several techniques that may be useful in this revision. Because these detection methods currently rely on genetic differentiation, researchers should consider the maintenance of reference specimens in cryogenic conditions which will permit future enzymatic and genotypic characterization of species. Such techniques can complement, and in some cases may surpass, our ability to distinguish closely related species based on morphology.
EDITORS NOTE:
On January 1, 1993, Science published UC Riverside findings that “sweetpotato whitefly strain B” is a distinct species not a strain. In the same issue, a news article described three unpublished studies supporting or challenging the UCR research. Correct identification of this devastating pest, renamed “silverleaf whitefly” by the UCR authors, is critical to nationwide, multimillion-dollar control efforts now underway. Here is an abbreviated version of the UCR findings and their implications.
Results of our work have bearing on the evaluation of insects when being considered for quarantine. Often insects that are known to exist in a region are not quarantined; this evaluation is made based upon morphological characteristics. Since we have documented that two morphologically similar insects are separate species, we suggest that the inclusion of additional discriminating techniques such as IEF and PCR, when available, be utilized. Additional species of Bemisia undoubtedly exist in the world, and like the sweetpotato and silverleaf whiteflies, these may be morphologically similar to species already known. Continued vigilance may prevent or retard the future movement of such pests into currently noninfested regions.
In the development of local, state, or regional management programs for the silverleaf whitefly, the distinction between the sweetpotato whitefly and the silverleaf whitefly is a crucial issue. At the center of these programs is the relevance of previously published work on the sweetpotato whitefly. Research on many aspects of the biology of the sweetpotato whitefly has been conducted throughout the world for over 100 years. These earlier studies are potentially of less significance in relation to the current problem because the pest whitefly is not the sweetpotato whitefly. Of particular concern is the host range of the new pest, its ability to transmit viruses known to be vectored by sweetpotato whitefly, and the applicability of previous research on management methods, including sampling plans, crop rotation and other cultural practices, biological control, and chemical pest management. An example of this is the ever-increasing host list of B. tabaci (reported to be as high as 600 species) which likely is the accumulation of host records for species other than the sweetpotato whitefly.
Proper systematic identification of whiteflies is the foundation for effective, long-term management strategies. For example, a traditional search for effective biological control agents would commence in the native home of the pest. For sweetpotato whitefly, the origin has been suggested as Middle Eastern, African, or the Orient. Although we do not know the place of origin of the silverleaf whitefly, data suggesting that it is from the same region as the sweetpotato whitefly are lacking. In a similar vein, the search for host plant resistance traits may be most effective where the silverleaf whitefly is native.
As noted above, the place of origin of the silverleaf whitefly currently is unknown, and its world distribution needs to be determined. With the recognition of a new species (and possibly a species complex) in the genus Bemisia, researchers should be less willing to assume that the whitefly in their region is B. tabaci. Until morphological features which can be used as distinguishing characters are found, methods such as the IEF and PCR amplified DNA may be required for species determination.
Whitefly identification with isoelectric focusing
Whiteflies normally are distinguished by microscopic examination of the immatures in the late stage of development. Typical characteristics used in this determination include number, size and location of pores, orifices, papillae, and setae. Unfortunately, at the present time we are unable to identify morphological characteristics that will distinguish the silverleaf whitefly, the insect discussed in the accompanying article, from the sweetpotato whitefly, which has been in California for nearly 70 years.
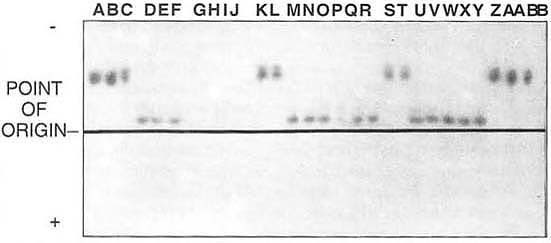
Biologists have developed other methods to distinguish closely related organisms. One that has become particularly useful for distinguishing the two species noted above is isoelectric focusing (IEF). It is a rapid assay that detects differences in enzymes which are indicative of genetic differences, and it can be done on individual whiteflies.
The IEF procedure is relatively easy to perform. We start by collecting whiteflies, the adults of which must arrive at the laboratory alive. If they die, their proteins begin to degrade and the IEF process no longer can be used. Therefore, most often we collect leaves infested with late-instar whitefly nymphs, bring these to the laboratory, and place the leaves in a Petri dish until whitefly adults emerge. The adults are collected and either analyzed immediately or stored in vials surrounded by liquid nitrogen (-270° C) until they are analyzed.
Once whiteflies have emerged and are collected (a process that can take several days) they are crushed individually onto a cellulose acetate membrane which has been soaked in a carrier solution to facilitate protein migration. The membrane is placed on a cooling plate that is kept at 4° C by a refrigerated antifreeze solution. The next step is to put a negative charge at one end of the membrane and a positive charge at the other end, which creates a charge gradient across the membrane. By subjecting the membrane to a timed series of voltages, the proteins migrate towards one of the poles and stop at a location specific for each enzyme. After the proteins have migrated, the membrane is stained with one of many different enzyme stains, and this allows us to visualize how far the proteins moved across the membrane. The IEF procedure requires about 3.5 hours for 120 individual whiteflies.
The results from one analysis of the enzyme phosoglucomutase is shown in the accompanying illustration. Protein variation between the two whitefly species is seen in the different migrational distances from their points of origin. Controls can be seen in lanes A, B, C, Z, AA, BB, which are from our silverleaf whitefly culture, and lanes D, E, F, W, X, Y, which are from our sweetpotato whitefly culture. Lanes K, L, S, and T were unknown whiteflies, now confirmed to be silverleaf whiteflies. Lanes M, N, O, Q, R, U, and V were unknown whiteflies, now confirmed to be sweetpotato whiteflies. T.P., C.F., T.B., A.C. and R.R.
Evidence for a new species of whitefly: UCR findings and implications

