


All Issues
Monterey pine forest made a remarkable recovery from pitch canker
Publication Information
California Agriculture 74(3):169-173. https://doi.org/10.3733/ca.2020a0019
Published online October 20, 2020
PDF | Citation | Permissions
NALT Keywords
Abstract
Monterey pine (Pinus radiata) is a species of limited distribution, with three native populations in California. In 1986, a disease known as pitch canker, caused by the fungus Fusarium circinatum, was identified as the cause of extensive mortality in planted Monterey pines in Santa Cruz County. Monitoring studies on the Monterey Peninsula documented rapid progression of the disease in the native forest during the 1990s, with most trees sustaining some level of infection. However, between 1999 and 2013, the severity of pitch canker stabilized, with many previously diseased trees then free of symptoms, and plots monitored between 2011 and 2015 documented a steady decline in the occurrence of new infections. Consequently, whereas pitch canker was once a conspicuous visual blight in the forest, by the end of the observation period, symptomatic trees had become a rarity. The arrested development of pitch canker is suggestive of a reduction in the frequency and duration of fog near the coast, which provides conditions necessary for the pathogen to establish infections.
Full text
Monterey pine (Pinus radiata D. Don) is native to California, where the distribution is limited to three populations on the Central Coast: Cambria in San Luis Obispo County, Monterey in Monterey County and Año Nuevo in Santa Cruz and San Mateo counties (Rogers 2002). As the dominant tree species throughout most of its native range, Monterey pine is of great ecological value, and a defining scenic feature in recreational areas such as Point Lobos State Natural Reserve and San Simeon State Park. Native populations are also an important genetic resource because Monterey pine is one of the most widely planted species in plantation forestry. Monterey pine plantations, which occupy over 10 million acres worldwide (Rogers 2004), make a particularly significant contribution to the economies of New Zealand, Chile and Australia.

Examining Monterey pines for evidence of wounds caused by twig beetles, a vector of the pitch canker pathogen. Researchers found the disease to be more severe near the coast than farther inland, suggesting the frequency and duration of fog may influence development of pitch canker.
The local and global importance of Monterey pine incentivizes maintaining the integrity of native populations, all of which are threatened by urbanization, and the associated need for fire suppression. In the absence of fire, growth of understory vegetation limits opportunities for pine seedlings to establish. Another threat to native populations is an exotic disease known as pitch canker, caused by the fungus Fusarium circinatum Nirenberg & O'Donnell.
Pitch canker was first described by Hepting and Roth (1946) in North Carolina, where the disease was discovered in a mixed-species plantation. Virginia pine (Pinus virginiana), shortleaf pine (P. echinata) and pitch pine (P. rigida) were all observed to be susceptible to pitch canker. Over the next several decades, pitch canker became a widespread problem in plantations and seed orchards in the southeastern United States (Dwinell et al. 1985). In 1986, pitch canker was identified as the cause of extensive mortality in Monterey pine in coastal California (McCain et al. 1987). Genetic evidence indicates the pathogen was moved to California from the infestation in the southeastern United States (Wikler and Gordon 2000), most likely on contaminated seed (Gordon et al. 2001).
Infections by the pitch canker fungus cause resinous lesions that girdle the affected branch, resulting in death of tissues distal to the infection. Infections on the trunk can cause decline and death of the entire tree. Trees weakened by pitch canker are attractive to native insects such as engraver beetles (Ips spp.), which increase the risk of mortality (Erbilgin et al. 2008).

Resinous lesion on a Monterey pine branch infected by the pitch canker pathogen, Fusarium circinatum. Removal of bark reveals discolored tissue killed by growth of the pathogen.

Native Monterey pine with typical early symptoms of pitch canker. Yellow-brown needles result from an infection that has girdled a branch proximal to the dying needles.

Monterey pine with multiples sites of pitch canker infection, resulting in brown needles and naked tips where branches have been girdled.

Monterey pine killed by the pitch canker pathogen, Fusarium circinatum. Tree death is associated with multiple infections on the trunk.
In California, severe impacts of pitch canker on Monterey pine were observed in off-site (i.e., outside the native range) plantings bordering State Highway 1 in Santa Cruz County as early as 1986 (McCain et al. 1987). This was consistent with the history of pitch canker in the southeastern United States, where the disease was problematic in managed stands and not in native forests. However, by 1994, pitch canker was confirmed in all three native populations of Monterey pine in California. Bishop pine (P. muricata) and knobcone pine (P. attenuata), two other closed-cone pine species native to California, have also been damaged by pitch canker (Gordon et al. 2001).
The goal of our research was to understand the factors that influence severity of pitch canker in the native Monterey pine forest. To this end, we recorded the incidence and severity of pitch canker over time, assessed the frequency of disease resistance among standing trees in the forest and quantified the abundance of insects that can serve as vectors of the pitch canker pathogen.
Monterey Peninsula research plots
Plots were established in the native forest on the Monterey Peninsula in 1996 (hereafter referred to as 1996 plots) and monitored for incidence and severity of disease through 2013. Forty plots were located west of Highway 1 in Monterey County, where the largest contiguous stands of native Monterey pine are found. Thirty-nine of the 40 plots were composed solely of Monterey pine. One plot had only bishop pines, and data from that plot were excluded from the analyses presented here. Of the remaining 39 plots, 18 were within 0.9 miles (1.5 kilometers) of the coast (= coastal plots) and 21 were located farther inland (= inland plots).
Each plot included 15 trees of at least 49 inches (125 centimeters) in height. Trees were rated for incidence and severity of pitch canker as described by Wikler et al. (2003). Each tree was categorized as having 0, 1 to 10, or > 10 symptomatic shoot tips. Trees were also placed in categories based on the number of cankers on the trunk of the tree: 0, 1 to 3, or > 3. Data on tip and trunk symptoms were combined to give each tree a disease severity rating, such that 1 to 10 tips counted as 1 point, over 10 tip infections counted as 2 points, 1 to 3 stem cankers counted as 1 point, and over 3 trunk cankers counted as 2 points. Thus, the minimum tree disease severity rating was 0, and the maximum was 4.
In 2011, six additional plots were established to monitor development of pitch canker in a cohort of younger trees (= 2011 plots). Each of the 2011 plots included 50 or 51 trees, with a DBH (diameter at breast height) ranging from 0.4 to 4.3 inches (1 to 11 centimeters). Following an initial assessment in September 2011, plots were evaluated three times per year in 2012 and 2013 (January, May and September), with additional assessments in January and July 2014, and January 2015. On each assessment date, the number of new pitch canker infections on each tree was recorded. New infections were distinguished from pre-existing infections based on symptoms, as described by McNee et al. (2002).
Previous work showed standing trees in the Monterey pine forest to vary in susceptibility to pitch canker, based on the length of lesions that developed at the site of inoculation (Storer et al. 1999). Research has also documented that infections by the pitch canker pathogen can enhance resistance to subsequent infection attempts, a phenomenon referred to as induced resistance (Bonello et al. 2001). To determine how this may have affected the abundance of resistant trees on the Monterey Peninsula, a selection of Monterey pines was inoculated in 2011 and the resulting lesions measured, as described by Gordon et al. (1998).
Each tree was inoculated on three branches by creating a wound approximately 0.06 inch (1.6 millimeters) in diameter that penetrated the bark in the inter-nodal area between terminal and penultimate branch whorls and depositing therein an aqueous suspension of approximately 250 spores of F. circinatum. Inoculum was prepared as described by Schmale and Gordon (2003). A total of 133 trees were located in six plots (separate from plots established for monitoring in the same year) in native stands on the Monterey Peninsula, with a minimum of 20 trees at each site.

A funnel trap baited with the pheromone pityol was used to trap twig beetles. The pitch canker pathogen was recovered from insects on eight of 14 sampling dates, at levels that ranged from 0.1% to 4%.
To assess the abundance of twig beetles (Pityophthorus spp.), which can serve as vectors for the pitch canker pathogen (Erbilgin et al. 2009; Sakamoto et al. 2007), funnel traps were deployed as described by Storer et al. (2004). Traps were baited with pityol, which is attractive specifically to P. setosus, one of the most common twig beetle species in the Monterey pine forest (Dallara et al. 2000). Traps were placed at six locations and sampled periodically from September 2011 through May 2013. At each sampling date, the number of twig beetles recovered was recorded, along with the percentage of insects carrying spores of the pitch canker pathogen, as described by Storer et al. (2004).
Disease severity leveled off
In the 3 years following establishment of plots on the Monterey Peninsula (1996 to 1999), the incidence and severity of pitch canker increased dramatically (Wikler et al. 2003). Furthermore, in a previous study of pitch canker in planted Monterey pines, only 2.2% of trees in heavily diseased plots remained free of infection, indicating that nearly all trees were susceptible to the disease (Storer et al. 2002). Consequently, it seemed likely that damage from pitch canker would continue to increase, with a potentially irreversible, negative impact on the forest. Contrary to this expectation, disease severity in the 1996 plots leveled off and changed very little over the next 14 years (fig. 1).

FIG. 1. The mean percentage of maximum severity of pitch canker in coastal (n = 18) and inland (n = 21) plots of Monterey pine on the Monterey Peninsula established in spring 1996 and evaluated at intervals until summer 2013.
In part, stabilization of the disease may be due to induced resistance, acting to reduce susceptibility of trees that sustained prior infections. Induced resistance may also help to explain a very low frequency of infection of trees in the 2011 plots. Assessments of 304 trees revealed a total of 27 and 63 new infections at 4 and 8 months after plots were established, respectively. Thereafter, the number of infections declined steadily, with a total of 14 recorded on the final assessment date in 2015. Over the 3 years that plots were monitored, 50% of trees (151/304) sustained no infections, and 22% (66/304) had only one (fig. 2).

FIG. 2. The number of Monterey pines sustaining between zero and eight infections by the pitch canker pathogen in the Monterey pine forest from September 2011 through January 2015.
If induced resistance was an important contributor to the reduction in the incidence and severity of pitch canker, most trees in the forest should be resistant to the disease. The results of the inoculation trial revealed a wide range of responses to inoculation, with mean lesion lengths (n = 3) ranging from 0.1 to 3.3 inches (2.5 to 85 millimeters). Lesions ≤ 0.6 inch (15 millimeters) in length are unlikely to girdle branches and hence no visible symptoms will develop. Therefore, lesion lengths below this threshold are indicative of resistance to pitch canker (Gordon et al. 1998). On this basis, 41.3% (55/133) of trees inoculated in 2011 were resistant, compared to 38.8% of Monterey pines inoculated with F. circinatum in 2002 (Gordon et al. 2011). This comparison suggests there was relatively little change in resistance of Monterey pines to pitch canker in the native forest on the Monterey Peninsula.
F. circinatum is a wound-infecting pathogen, and, in California, wounds created by twig beetles (Pityophthorus spp.) can serve as infection courts (Sakamoto et al. 2007; Storer et al. 2004). The number of twig beetles recovered from traps in the Monterey pine forest between September 2011 and May 2013 ranged from 37 to 677. There was no apparent trend across this time interval (fig. 3). The pitch canker pathogen was recovered from insects on eight of 14 sampling dates, at levels that ranged from 0.1% to 4%.
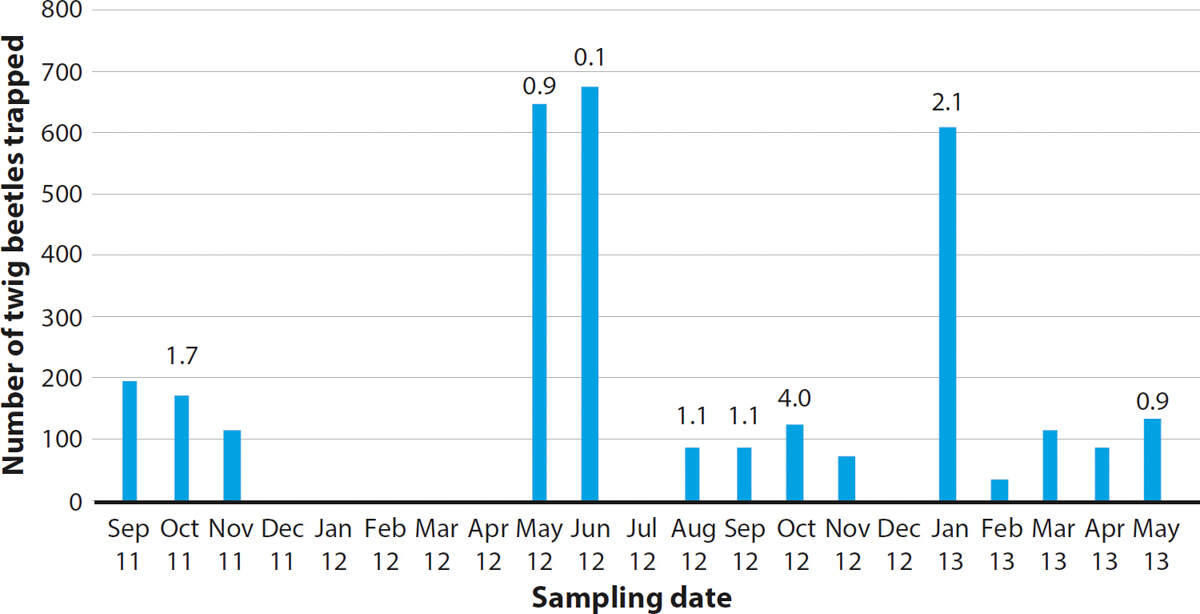
FIG. 3. The number of twig beetles recovered from traps baited with the pheromone pityol on the dates (month/year) indicated. The height of each bar represents the total for traps placed at six locations in the Monterey pine forest. The number above the bar is the percentage of beetles from which Fusarium circinatum was isolated. The absence of a number indicates that the pathogen was not recovered from any twig beetles on that date.
Two of the sites included in the present study were also sampled in 1999. In that year, recovery of F. circinatum from trapped twig beetles, across three sampling dates at both locations, ranged from 0.9% to 17% (Storer et al. 2004). Thus although twig beetles remained abundant in the forest, fewer of them were carrying the pathogen. Whether this was a consequence of fewer trees with active infections, and hence fewer spores being produced, or it was among the causal factors reducing the incidence of pitch canker cannot be determined.
Environmental effects on disease
Another factor that could influence development of pitch canker is the frequency with which environmental conditions are conducive to infections. One indication of an environmental effect on the infection process was greater disease severity in coastal plots than in inland plots on the Monterey Peninsula. This differential in disease severity changed very little over a span of 17 years (fig. 1) and may be due, in part, to greater frequency and duration of fog in the coastal plots. Consistent with this interpretation are data showing a higher rate of infection in trees maintained at 100% relative humidity than in trees exposed to a lower relative humidity (Sakamoto et al. 2007).
This is relevant to the arrested development of pitch canker on the Monterey Peninsula if the frequency and duration of fog events decreased along the Central Coast, which would have made moisture required by the pathogen less available, thereby reducing the risk of infection. Indeed, in the typical summer fog seasons of 2014 and 2015, particularly during the normally foggy month of August, data collected by D. Fernandez (unpublished data) indicated significantly lower fog levels than in the preceding 4 years. His two data sites were near sea level and within 20 kilometers of our Monterey pine study sites on the Monterey Peninsula.
Over a period of 16 years, a remarkable recovery from pitch canker was evident in the Monterey pine forest on the Monterey Peninsula. At the start of this interval, the disease was a highly visible blight on the landscape, and by the end, few symptomatic trees could be found in most parts of the forest. Although induced resistance can help to explain the recovery of previously symptomatic trees, most standing trees in 2011 appeared to be susceptible to the disease, and insect vectors carrying spores of the pathogen could still be recovered in the forest. However, infections require relatively warm conditions when moisture is present (Inman et al. 2008). On the Monterey Peninsula, this requirement was met through high humidity and deposition associated with summer fog. It may be that a reduction in frequency with which moisture was made available through fog played a key role in the diminished impact of pitch canker.
Observations suggest that a dramatic decline in disease severity has also occurred among planted pines in Monterey and Santa Cruz counties. Less information is available on development of pitch canker in other locations, but fewer and shorter intervals of fog may well have an effect similar to what we documented on the Monterey Peninsula.
Monterey pine forest made a remarkable recovery from pitch canker

