


All Issues
Possible impacts of rising CO2 on crop water use efficiency and food security
Publication Information
California Agriculture 72(3):155-158. https://doi.org/10.3733/ca.2018a0028
Published online September 19, 2018
PDF | Citation | Permissions
NALT Keywords
Summary
Understanding the mechanisms involved in plants' response to rising CO2 levels may lead to the development of crop plant varieties better adapted to future drought conditions.
Full text
California and the rest of the world are experiencing major changes in the availability of water and the concentration of atmospheric carbon dioxide (CO2). Water and CO2 — along with energy from the sun — are the inputs to photosynthesis, the basis of plant growth and food production. Elevated CO2 can influence the water use efficiency and yield of crop plants. A clearer understanding of the mechanisms involved in those responses may lead to the development of crop plant varieties better adapted to expected future water and CO2 conditions.
In recent years, the United States has experienced an increased frequency of heat waves and drought and related loss of crops (Lesk et al. 2016). In 2014, drought cost California an estimated $2.2 billion in lost agricultural production (Howitt et al. 2015). In California and elsewhere, the changing climate, population growth and mandates for sustainable management of surface water and groundwater are likely to make water for irrigation more scarce in the future.
From 1950 to 2000, global atmospheric CO2 concentration rose 33%. It continues to climb (fig. 1) and is a major cause of the continuing rise in global temperature (Victor et al. 2014). Average temperatures in California have risen 2.5°F since 1880, and this rate is expected to increase over the next century due to emissions of greenhouse gases (Pathak et al. 2018). Among other impacts, the rising temperature is expected to reduce the winter snowpack, effectively reducing California's water storage capacity and useful water supply (PPIC 2016).

FIG. 1. The continuing steep increase in the atmospheric concentration of CO2. Used with permission from Dr. Ralph F. Keeling, UC San Diego's Scripps Institution of Oceanography CO2 program.
CO2 and stomatal pore apertures
Research in our laboratory investigates the mechanisms by which CO2 elevation and drought cause closing of leaf stomata. Plant stomata are pores on leaf surfaces that enable the intake of CO2 and the release of water vapor. Over 90% of the water lost by plants through evapotranspiration is released via stomata (Hetherington and Woodward 2003).
Each stoma is surrounded by two specialized guard cells that open and close the central gas exchange pore (fig. 2). CO2 closes stomatal pores, by entering the guard cells, where it is converted to bicarbonate. The increased concentration of bicarbonate is thought to activate molecular switches called protein kinases, which in turn open channels that release ions from the guard cells (Hu et al. 2010). The release of ions reduces the turgor pressure of the guard cells, in effect deflating them and closing the stomatal pore (fig. 2) (Engineer et al. 2016). This process is part of the daily cycle of plant photosynthesis and respiration (in which the glucose generated by photosynthesis provides energy for the plant). After the sun sets and plants can no longer photosynthesize, respiration increases the CO2 concentration inside plant leaves, causing the stomata to close.
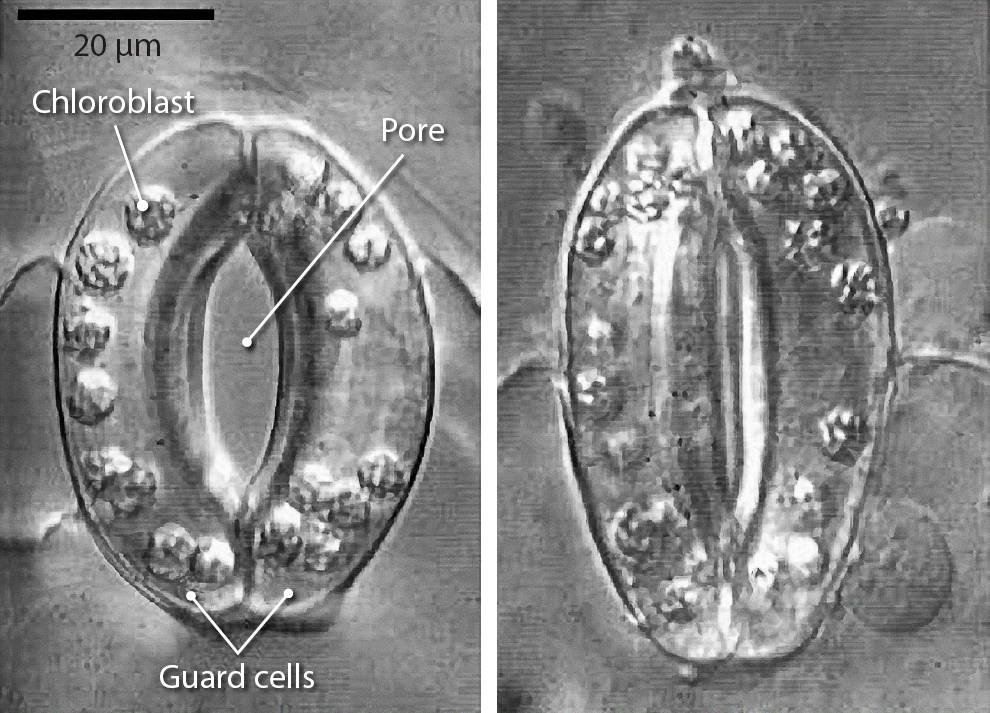
FIG. 2. Light microscopy of plant stomatal pores in the epidermis of leaves in open, left, and closed, right, conformations. Stomatal pores are embedded in the epidermis of the leaf, regulating water loss from the leaf and CO2 uptake from the atmosphere. The two guard cells surround the central pore through which water is lost by plants. Image reproduced from Plant Physiology, 2nd Edition, p. 523, edited by Taiz and Zeiger. By permission of Oxford University Press, USA.
Potential for water use efficiency gains
Optimization of CO2-controlled stomatal closing could lead to development of plant varieties for water-scarce environments that lose substantially less water per carbon fixed by photosynthesis. Our research has identified two enzymes, ß-carbonic anhydrases ßCA1 and ßCA4, which, when overexpressed in guard cells in Arabidopsis thaliana, enhance CO2-regulation of stomatal closing while maintaining functional drought and light responses. This resulted in an average of approximately 40% less water loss by the plant per carbon fixed via photosynthesis in the laboratory (Hu et al. 2010). In another example, overexpression of epidermal patterning factor 1 and 2 (EPF1 and EPF2) in barley (Hordeum vulgare), HvEPF1, reduced the density of stomata without affecting grain yield. The reduced stomatal pore apertures (Hu et al. 2010) and density (Hughes et al. 2017) resulted in improved water use efficiency and drought resistance, pointing to applications for water-scarce environments (Hu et al. 2010; Hughes et al. 2017). Since plants draw water out of the soil and accelerate deep soil drying, reduced stomatal apertures and density can serve to maintain more water in the soil, thus reducing damaging effects of longer-lasting droughts. The effects of changing stomatal physiology have yet to be evaluated in crop plants grown in the field, but these laboratory studies illustrate the potential for advances.
Related potential for increased yield
While stomatal closure at elevated CO2 or lower stomatal density may improve water use efficiency in crops in water limited environments, the opposite manipulations could be effective in the future to maximize yield in regions with ample water and precipitation. In regions where water and soil nutrients are sufficient, increasing crop productivity is a priority. In these regions, developing plants that respond less strongly to CO2 in their stomatal closing response — thus enabling enhanced CO2 intake by leaves — may result in increased yields.
There are several distinct forms of photosynthesis that plants use. Two common forms are C3 photosynthesis in plants such as wheat, beans, tomatoes, rice, soybeans and all trees; and C4 photosynthesis in plants such as maize, quinoa and sorghum. C3 crops are also grown in temperate regions of the world with ample supplies of water. C3 photosynthesis is less efficient than C4 photosynthesis, and C3 photosynthesis is not yet working at maximum capacity at present atmospheric CO2 concentrations. For C3 plants growing with sufficient water and nutrients, maintaining more open stomata at our present high atmospheric CO2 levels may be beneficial for yields depending on the crop type (Mohammed 2013).

For C3 crops — like wheat, beans, tomatoes, rice, soybeans and all trees — maintaining more open stomata in a high-CO2 environment with sufficient water and nutrients may in some cases improve yields.

For C4 crops like maize, quinoa and sorghum, reducing stomatal apertures while enabling optimal CO2 intake for C4 photosynthesis may increase water use efficiency.
An advanced understanding of the mechanisms and genes that mediate CO2 regulation of stomata could become valuable for better adjusting C3 plants in light of the steeply increasing atmospheric CO2. For most C4 crops, photosynthesis is generally already working at maximum capacity, due to the increased atmospheric CO2 concentration; such crops would thus instead be candidates for the manipulations described above that aim to increase water use efficiency by reducing stomatal apertures while enabling optimal CO2 intake for C4 photosynthesis.
Stomatal density, heat tolerance
One important potential complication: elevated CO2 also tends to reduce the density of stomata on leaves of some plant species (Woodward 1987). This effect, together with CO2-induced reduction in stomatal apertures, can reduce a plant's capability to cool itself because of reduced evaporation of water from leaves. This may increase susceptibility to heat stress (Engineer et al. 2016), which in turn can affect water use and yield.
In a recent study of soybeans exposed to elevated CO2, the CO2-induced reduction in stomatal pore density and aperture resulted in increased leaf temperature and a net increase in water demand, compared to soybean plants grown in environments with lower CO2 concentrations (Gray et al. 2016).
Thus, the effects of stomatal manipulations will need to be tested in the field and their benefit may depend on the heat response of individual crops.
Future outlook
Present research advances have led to an initial understanding of mechanisms by which plants sense CO2 concentration and transduce the CO2 signal to regulate water loss. This research is enabling the development of plants with increased water use efficiency and potentially improved yields, depending on growing conditions. However, many of the proteins and genes that make up the CO2 signal transduction network still remain unknown, and how these network components interact within the network is largely not understood at present. Uncovering the missing links in CO2 signal transduction is the focus of work in our laboratory and in other laboratories. Future research will be required to fully uncover all genes and proteins that cause CO2 regulation of stomatal pores. This research promises to provide not only an in-depth understanding of how plants respond to CO2 concentration changes, but also will provide advanced tools for breeding aimed at optimizing those responses to water and CO2 and climate conditions. This research is needed to help safeguard the California and U.S. agricultural economy and the food, feed and fuel it provides.
Possible impacts of rising CO2 on crop water use efficiency and food security

