


All Issues
Irrigation method does not affect wild bee pollinators of hybrid sunflower
Publication Information
California Agriculture 71(1):35-40. https://doi.org/10.3733/ca.2016a0017
Published online September 13, 2016
NALT Keywords
Abstract
Irrigation method has the potential to directly or indirectly influence populations of wild bee crop pollinators nesting and foraging in irrigated crop fields. The majority of wild bee species nest in the ground, and their nests may be susceptible to flooding. In addition, their pollination of crops can be influenced by nectar quality and quantity, which are related to water availability. To determine whether different irrigation methods affect crop pollinators, we compared the number of ground-nesting bees nesting and foraging in drip- and furrow-irrigated hybrid sunflower fields in the Sacramento Valley. We found that irrigation method did not impact wild bee nesting rates or foraging bee abundance or bee species richness. These findings suggest that changing from furrow irrigation to drip irrigation to conserve water likely will not alter hybrid sunflower crop pollination.
Full text
Irrigation practices and water use efficiency are increasingly scrutinized by growers. Irrigated agriculture accounts for 80% of human-related water use in California (DWR 2013). In periods of drought, growers adopt water-saving irrigation practices at higher rates (Schuck et al. 2005). Drip irrigation, introduced to California in 1969, delivers water directly to the plant root zone, thus improving water efficiency; it is now used in approximately 40% of all irrigated fields (Taylor et al. 2014). Increases in irrigation efficiency can improve yield (Tilman et al. 2002; Wallace 2000), which is another reason growers may consider switching to drip irrigation. However, changes in irrigation practices may negatively impact other factors that determine crop success, such as pollination.
Wild bees are the most effective and abundant crop pollinators (Garibaldi et al. 2013). The majority of wild bees excavate nests beneath the soil (known as ground nesters). Irrigation has the potential to saturate nests, possibly drowning bee larvae and adults. It could also indirectly impact crop pollinators by affecting their foraging choices. Bee foraging decisions are often related to floral reward, namely nectar quantity and quality (Roubik and Buchmann 1984; Stone 1994). Nectar production is related to water availability; increased water leads to higher nectar volume expressed (e.g., Petanidou et al. 1999). Thus, an irrigation method that delivers more water, such as furrow, could make fields more attractive to wild bee pollinators, thereby increasing potential yields.

Findings from a study of Sacramento Valley hybrid sunflower fields suggest that drip irrigation does not have a negative effect on native bee crop pollinators.
We compared the number of bees nesting and foraging in conventionally managed hybrid sunflower (Helianthus annuus) fields that were either furrow or drip irrigated. We predicted that drip-irrigated fields would support higher numbers of nesting bees, but that more bees would forage in furrow-irrigated fields, due to indirect effects.

The stages of furrow irrigation, also known as flood irrigation: during irrigation (A), following saturation (B) and after water applied to the field has dried (C). Drip-irrigated fields lack the cracking found in furrows of flood-irrigated fields; the soil surface appears dry, even during irrigation events (D).
We also examined whether irrigation had the same effect on different bee groups. Sunflower is visited by both specialist and generalist ground-nesting native bee pollinators that nest within crop fields (Hurd et al. 1980; Kim et al. 2006; Sardiñas et al. 2016). Generalist bees visit a variety of plant species, whereas specialists collect only sunflower pollen to provision their nests. While both of these types of pollinators could be susceptible to irrigation methods, sunflower specialists are more tied to the crop and could experience potential negative effects of irrigation more strongly.

Nest entrances of the ground-nesting sunflower specialist bee, Diadasia enavata (A). Sunflower is visited by both specialist and generalist pollinators, including the generalist Halictus ligatus (B, arrows) and the specialist Diadasia enavata (B).
Study design
We sampled five drip- and five furrow-irrigated sites in 2013 during the summer months at peak sunflower bloom (July to August). Site types were paired by bloom time, sunflower variety and landscape context (e.g., percent natural habitat within 1 km) to reduce extraneous factors that could contribute to differences in nesting and foraging patterns observed. All fields were located in Yolo County, California.
Bee sampling
We used 1.96-square-foot emergence traps (BugDorm, MegaView Science, Taiwan) to sample nesting bees (Sardiñas and Kremen 2014; Sardiñas et al. 2016). The traps have open bottoms to allow nesting bees to leave their nest. However, when they emerge, they are funneled to the top of the trap and into a kill jar. We placed the traps at dusk, after bees had returned to their nests, and weighted down the edges with soil to prevent bees from entering or exiting the trap. There were 20 traps in each field along two parallel 328-foot transects that ran into each field (fig. 1). Traps were 32.8 feet apart. Approximately 20 hours later, we removed all bees from apical kill jars (which were filled with soapy water).

Fig. 1. Emergence traps (A) were used to collect bees nesting in sunflower fields and were placed along two parallel transects (B) running 328 feet into the fields. Transects were located 164 feet from field edges and 328 feet apart. Ten traps (white boxes, B), 32.8 feet apart, were placed along each transect.
The day following emergence trap sampling, we netted foraging bees visiting sunflowers for 30 minutes along each of the two transects. We set emergence traps only if weather conditions the following day were predicted to be ideal for netting: temperature > 64°F, wind speed < 5.5 mph and low cloud cover (clear skies).
All bees were pinned, then identified by Dr. Robbin Thorp, professor emeritus, UC Davis Department of Entomology.
They are currently housed in UC Berkeley's Essig Museum of Invertebrate Zoology.
Vegetation
To determine whether vegetative factors influenced bee abundance or species richness, we estimated percentage sunflower bloom, stem density (count of all sunflower stems), weed density (count of all individual weeds) and weed bloom. Sunflower bloom and stem density were correlated, as were weed density and weed bloom, which allowed us to use only one metric for each category in our analyses.
Statistical analyses
We examined the effect of irrigation method on the abundance of nesting bees captured in emergence traps and foraging bees netted at blooms using a generalized linear model with a Poisson distribution in the R package lme4 (Bates et al. 2015). Independent variables were irrigation type, stem density and weed density. Site was a random effect. We repeated this analysis for species richness, which was calculated using the R package vegan (Oksanen et al. 2013). We included only female bees in our analyses of nesting rates, as male bees do not excavate nests (Kim et al. 2006).
Bee nesting and foraging counts
We collected 42 bees from six species nesting within fields and 735 bees from 14 species foraging on sunflower blooms (table 1). All of the species we collected nesting were also found foraging. The two most abundant species nesting in sunflower fields were the sunflower specialist Melissodes agilis and the generalist sweat bee Lasioglossum incompletum. These bees were among the most abundant bee species found foraging. One other species of sweat bee (Halictus ligatus) and three other sunflower specialist bees (Diadasia enavata, M. robustior and Svastra obliqua expurgata) foraged in high numbers, yet were not detected nesting within fields.

Bee response to irrigation method
We did not find a difference in the abundance of bees nesting in drip- versus furrow-irrigated fields (fig. 2A; z = 0.29, P = 0.77). Similarly, the species richness of nesting bees did not vary with irrigation type (fig. 2C; z = −0.40, P = 0.68). Sunflower stem density (z = 0.71, P = 0.48) and weed density (z = −0.15, P = 0.85) did not impact nesting rates (data not shown), which is not surprising given that we attempted to control for variability in bloom by sampling at peak bloom in all fields (> 90% of stems in bloom).
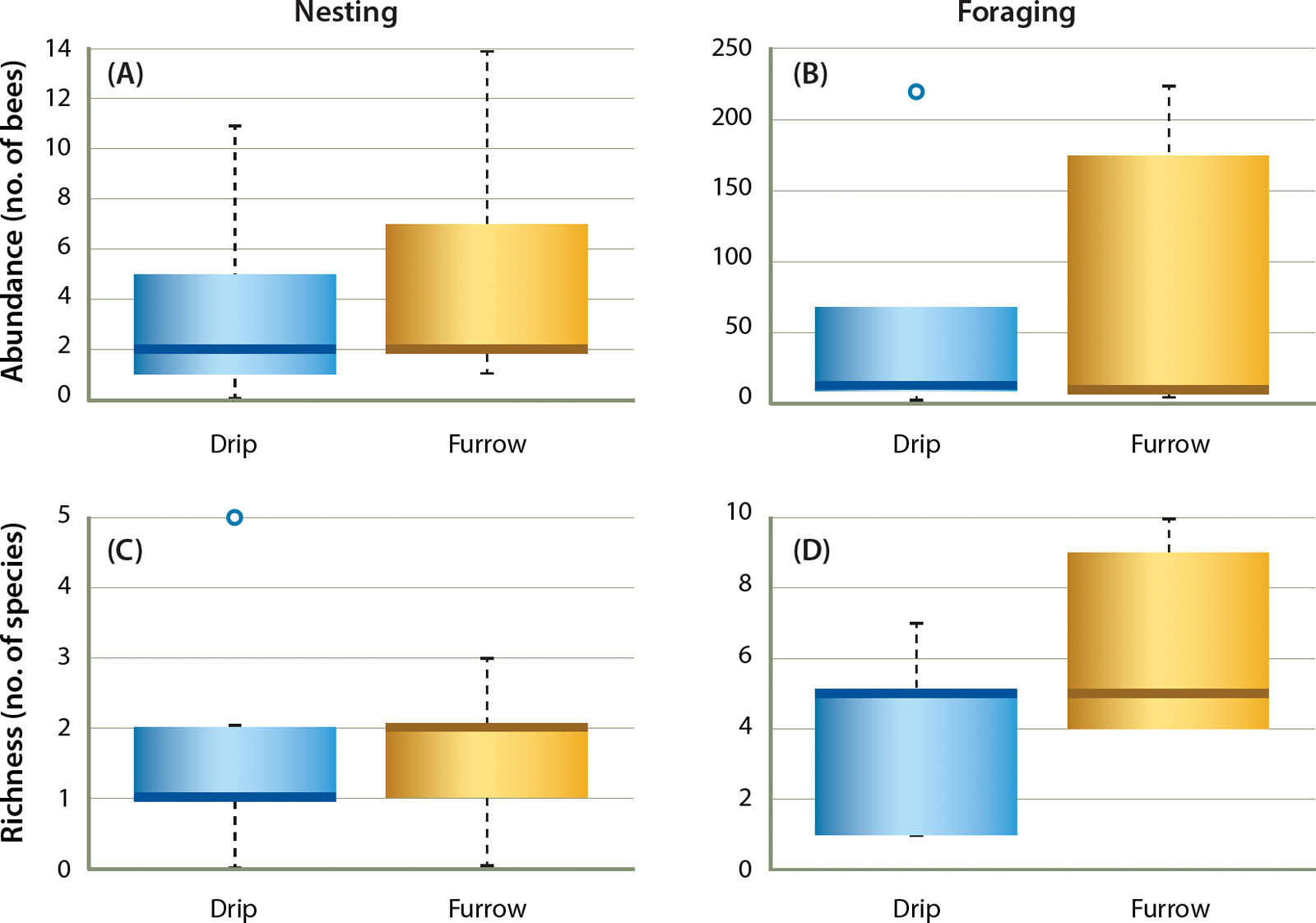
Fig. 2. Irrigation method did not affect the abundance or species richness of nesting bees (A, C) or foraging bees (B, D) in sunflower fields. Boxes are upper and lower quartiles, dark bar is the mean, whiskers show the maximum and minimum values, and points are outliers.
As with nesting rates, the abundance (fig. 2B; z = 0.12, P = 0.89) and species richness (fig. 2D; z = 1.60, P = 0.11) of native bees actively foraging on sunflower was unaffected by irrigation type. Sites that were sampled at the same time appeared to contain similar numbers of foraging bees (fig. 3) except for sites D3 and F3, where the drip-irrigated site contained almost twice as many foraging bees. However, we were unable to assess the effect of sampling date in our analyses as each site was sampled only once.

Fig. 3. Foraging bee abundance in site pairs (1–5), which were drip- (D) and furrow- (F) irrigated sunflower fields that had the same variety and bloom time.
Study sample size
This study was conducted during a single year; therefore, the results reflect nesting and foraging during this one sampling season. Our sample size may not have been large enough to detect small differences in nesting rates. While the strength of the nesting results indicates that sunflower bee nesting is likely not linked to irrigation method, additional evidence from future studies could help confirm this conclusion. We collected numbers and species of bees in our netted sample that were similar to those in other studies in sunflower in our study region (Greenleaf and Kremen 2006a; Sardiñas and Kremen 2015); this similarity suggests our findings on the relationship of foraging bees to irrigation type may be robust to the effects of small study size.
Soil moisture conditions
Soil moisture has been shown to positively affect nesting (Julier and Roulston 2009; Xie et al. 2013); therefore, irrigation may help make fields attractive nest site locations for crop pollinators. Soil moisture, however, may be correlated to a number of other conditions, including soil compaction (Xie et al. 2013). In the sunflower study system, generalist wild bees have been found to nest both within crop fields as well as along un-irrigated field edges (Sardiñas et al. 2016). Soil moisture may not exert as strong of effects as other characteristics that affect nest site selection and nesting success of generalists, while sunflower specialists may be better adapted to the within field conditions where sunflowers are grown.
Bees’ adaptation to inundation
The most abundant foraging and nesting sunflower specialist species, M. agilis, has been recorded nesting between irrigation furrows in crop fields since the early 1980s (Parker et al. 1981). The cells in their nests are lined with wax, which may have some hydrophobic properties (Cane 1981). Water-resistant wax linings have been recorded in the nests of other bee species (e.g., Rust et al. 2004).
Species whose nests are not regularly exposed to wet conditions may be able to withstand extreme conditions, such as flooding from a hurricane (Cane 1997), although some species may have local nesting aggregations wiped out by similar events if the soil structure is compromised, for example by mud slides (Fellendorf et al. 2004). The ability to withstand irrigation or natural saturation events has not been recorded for most of the bees in this study; however, the bees’ presence in regularly irrigated fields indicates that irrigation may not be a factor that significantly limits or disrupts their nesting activity.
Which irrigation method is best?
Although drip irrigation is often considerably more expensive than furrow irrigation, there are numerous benefits other than water use efficiency associated with drip irrigation, including disease management and the ability to irrigate oddly shaped or uneven fields (Shock 2013). Drip irrigation, especially subsurface drip, can reduce the total amount of acre-feet applied because it reduces evaporation (Ayars et al. 2015). Over 85% of processing tomato fields in California have been converted to drip irrigation systems, which has increased yields without compromising crop quality (Taylor et al. 2014). Sunflower is often rotated into fields that contained tomato the year prior because the two crops have similar row spacing. Growers leave the drip tape down (H. Sardiñas, personal observation), maximizing their investment through reuse of the drip tape.
Water efficiency is especially important in California's Central Valley, where climate change is expected to increase temperatures 2°F to 3.6°F by 2050 and the frequency, intensity and duration of summer heat waves are expected to increase (Jackson et al. 2011). The current drought is driving up the cost of water and limiting water access, leading growers to increase well drilling to obtain groundwater (Daniel Munk, UC Cooperative Extension, personal communication). Wells were expected to account for 53% of all irrigation water in 2015; however, increased rates of pumping caused by the prolonged drought has caused the water level to drop below the depth of many wells (Howitt et al. 2014); this excess pumping is also leading to land subsidence.
Although this study was conducted in hybrid sunflower fields, the irrigation methods applied are typical of those used in row crops throughout the Central Valley. The generalist sweat bees that nest and forage on sunflower are among the most common crop pollinators in the region, and pollinate a variety of crops from watermelon to tomato (Greenleaf and Kremen 2006b; Morandin and Kremen 2013). We would therefore expect our findings to apply to a number of different annual crop types.
The combined efficiency benefits and lack of negative effects on native bee crop pollinators indicate that drip irrigation is a viable method to combat the drought without compromising crop pollination from bees nesting within crop fields.

In 2013, researchers sampled five drip- and five furrow-irrigated sites in Yolo County at peak sunflower bloom (July to August).
Irrigation method does not affect wild bee pollinators of hybrid sunflower

