


All Issues
Low-toxicity baits control ants in citrus orchards and grape vineyards
Publication Information
California Agriculture 58(4):213-217. https://doi.org/10.3733/ca.v058n04p213
Published October 01, 2004
PDF | Citation | Permissions
Abstract
Effective ant control is critical for controlling honeydew-secreting homopteran agricultural pests such as whitefly and mealybug. Low-toxicity ant baits may more effectively control ants than the broad-spectrum insecticides currently used in California vineyards and citrus orchards. This study focused on developing effective ant baits for use in bait stations to control field ant and Argentine ant, which aggressively tend homopteran pests. In the Coachella Valley, field ant is associated with the vine mealybug, a destructive nonnative pest. We conducted preference experiments for various commercially available ant baits and a bait formulated with anchovy plus imidacloprid. Field ant preferred the anchovy baits above all others tested, and in field trials the anchovy bait with 0.005% imidacloprid significantly reduced foraging activity. Argentine ant is the primary ant pest in vineyards and citrus orchards of California's nondesert growing regions. We tested the efficacy of several chemical bait treatments, all of which significantly lowered Argentine ant populations.
Full text
Biological control programs aimed at suppressing honeydew-secreting pests on citrus and grape must include ant control in order to optimize the effectiveness of natural enemies. Honeydew provides a stable food source for several species of ants (Beattie 1985). To protect this food source, ants will disturb or kill predators and parasitoids (Gullan 1997).

Field ants tend a vine mealybug colony, which is infesting the current season's girdling wound.
Citrus is often attacked by honeydew-producing homopterans such as soft scale (Coccus hesperidium L.), citrus mealybug (Planococcus citri Risso) and wooly whitefly (Aleurothrixus floccossus [Maskell]). Moreno et al. (1987) showed that the natural enemies of these pests provided more effective control when the Argentine ant (Linepithema humile [Mayr]) was absent. Itioka and Inoue (1996) demonstrated that ladybird beetles (Coccinellidae) and green lacewings (Chrysopidae) reduced a population of the mealybug (Pseudococcus citriculus Green) on Satsuma orange by 94% when the ant (Lasius niger L.) was controlled.
In California, three mealybug species — grape mealybug (Pseudococcus maritimus [Ehrhorn]), longtailed mealybug (Pseudococcus longispinus [Targioni-Tozzetti]) and obscure mealybug (Pseudococcus viberni Maskell) — commonly attack table, raisin and wine grape varieties. Predators such as lady beetles (Cryptolaemus sp.) and lacewings attack each of these species, and up to five species of parasitoids are known to attack the grape and longtailed mealybugs. In addition, the vine mealybug (Planococcus ficus [Signoret]) is a recent arrival into California, first reported on table grapes in the Coachella Valley (Riverside County) in 1994. Since its arrival, the vine mealybug has been reported on raisin and table grapes in the San Joaquin Valley and on wine grapes in Santa Barbara, San Luis Obispo, Sonoma and Napa counties, for a total of about 30,000 acres statewide. Although little research has been conducted on the interaction between ants and the natural enemies of these mealybugs on grape, ant suppression is thought to play a critical role in their control.
In the Coachella Valley, the field ant (Formica perpilosa [Wheeler]) is the predominant ant species associated with vine mealybug. The acrobat ant (Crematogaster sp.) is also present but to a lesser extent. Two fire ants, the southern fire ant (Solenopsis xyloni [Wheeler]) and the thief ant (Solenopsis molesta [Say]), are also found; however, their interaction with vine mealybug is not fully understood. The native gray ant (Formica aerata [Francoeur]) is similar to the field ant and is the primary ant associated with vine mealybug in the San Joaquin Valley and San Luis Obispo County. The Argentine ant, as well as being a primary ant pest in citrus orchards of the coastal growing regions, is also the primary ant pest associated with vine mealybug in the wine vineyards of Sonoma and Napa counties.

A choice-test arena (without lid) is used to determine the bait preferences of field ants.
Limitations of ant control methods
Current strategies to control ants in orchards generally include the application of residual insecticides, such as chlorpyrifos. Chlorpyrifos applied to the trunk and ground beneath a citrus tree provides a repellent barrier and kills foraging workers that come into contact with the insecticide. However, only limited control can be achieved because the queen or queens and the vast majority of workers in the nest are not affected. The effectiveness of chlorpyrifos is also limited by high temperature, irrigation and ground cover (Rust et al. 2000). When using this method, citrus trees must be skirt-pruned in order to prevent the ants from using alternative routes into the canopy. Trunk banding with an insecticide-impregnated paper or a sticky material is an effective method to prevent ants from foraging in the canopy. However, trunk banding is labor intensive and has not been adopted by the majority of growers.
Chlorpyrifos is also the primary method to control ants in vineyards, with similar limitations. Modifications to application techniques, which concentrate the spray to where the ants are located, have improved its effectiveness. Phillips and Sherk (1991) improved the efficacy of chlorpyrifos to approximately 8 weeks by using a hand-held, compressed-air sprayer and concentrating the application to the trunk and trunk-soil interface. Klotz et al. (2003) improved on this approach and designed a spray rig mounted on an all-terrain vehicle, which delivered the insecticide to a 2-foot-wide strip of soil beneath the grapevines where field ant nests. This method provided control of field ant for approximately 9 weeks.
Development of baits and stations
Toxic baits may offer a more effective method for controlling ants in orchards and vineyards. The recruitment and food-sharing behavior of ants can be exploited in order to spread a toxicant through the colony. Ant baits generally contain three components — a matrix or carrier, feeding stimulant and toxicant. Each of these components plays a critical role in the bait's attractiveness, which makes developing an effective ant bait challenging. For example, the particle size of the carrier affects the rate of collection (Hooper-Bui and Rust 2000); typically, larger ants prefer a large particle size. The optimal percentages of carbohydrate, protein and fat in the feeding stimulant may vary according to the species of ant or the nutritional requirements of a colony. Also, the toxicant must not deter feeding and must be slow acting so as not to impede recruitment and food sharing (Rust et al. 2000).
Bait delivered in stations minimizes environmental exposure to the toxicant, but developing an effective and economically feasible bait station also presents a challenge. The biology of the ant and its foraging behavior must be understood when considering the volume of bait that a station should contain and the number of stations needed per acre. For example, population densities of Argentine ant can be quite large. Markin (1967) conducted experiments in a citrus orchard in San Diego County and estimated that between 50,000 and 600,000 Argentine ants could ascend an individual citrus tree in a single day. Also, Argentine ants do not act aggressively toward nearby nests. Instead, they form a cooperative network of colonies throughout an area that represent one giant “supercolony.” The biology and foraging behavior of field ants are in sharp contrast to that of the Argentine ant; field ants do not form large colonies and are territorial. We are in the initial stages of bait development and have made progress in finding preferred feeding stimulants and toxicants for field ant and Argentine ant.
Field ant baits and efficacy
Bait tests
Under field conditions, we tested five commercially available baits to control field ant: 1.0% hydra-methylnon (Maxforce, Maxforce Insect Control Systems, Oakland, Calif.), 1.0% hydramethylnon (Maxforce, fine granular), 0.5% pyriproxyfen (Esteem, Valent U.S.A., Walnut Creek, Calif.), 5.0% orthorboric acid (Niban, Rockford, Tenn.) and 0.5% pyriproxyfen (Combat, Combat Insect Control, Pleasanton, Calif.). In addition, we tested a granular bait formulated with anchovy and 20-30 mesh corn grit developed in the laboratory of Dr. Michael Rust. The anchovy bait was tested with 0.005% and 0.05% imidacloprid.
These active ingredients generally have lower acute toxicity (LD50) than commonly used broad-spectrum organophosphate insecticides. However, what makes these “low-toxicity” baits is their dose. The anchovy bait formulated with imidacloprid is unique; the authors are not aware of any such ant baits on the market.
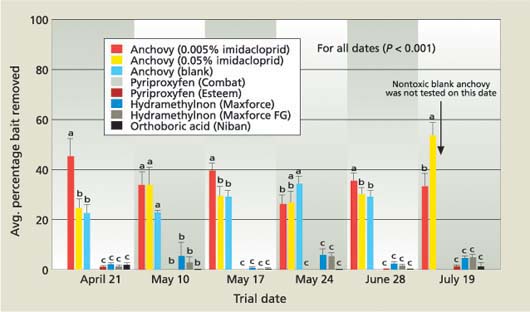
Fig. 1. Mean percentage bait removed (± SEM) by field ant in choice-test arenas placed at the base of ant-infested grapevines (n = 10). Ants were allowed to forage freely for 24 hours. On May 24, pyriproxyfen (Combat) replaced pyriproxyfen (Esteem) in each of the choice-test arenas.
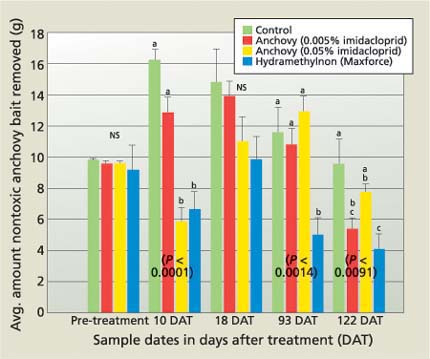
Fig. 2. On each sample date, population densities before and after a single insecticide treatment were monitored by allowing field ant to forage on about 20 grams of anchovy bait (nontoxic blank) placed at the base of four grapevines per plot and calculating the amount removed after 24 hours.
The bait preference test was a completely randomized design conducted in a 20-acre block of ‘Superior Seedless’ grapes. A visual inspection of the vineyard indicated that it was heavily infested with field ants. Approximately 6 grams of each bait was placed in a 25-milliliter (ml.) plastic vial and then randomly assigned to a position in a choice-test arena. The arenas were constructed from aluminum pans (12 inches in diameter by 2 inches deep) with four equidistantly spaced holes located on the side of the pan flush with the bottom. Glass tubing (3/8 inch inside diameter and about 4 inches long) was inserted into each hole, so that ants entering the arena would emerge from the glass tubing at the center. Twelve small weigh boats (1-inch-by-1-inch-by-0.5-inch plastic dishes used to weigh powder and liquid compounds) were glued to the floor of the pan equidistantly spaced from one another along the inside perimeter. The weigh boats were modified so that the vials could be laid flat within them and locked in place. In the field, each arena was covered with a piece of plywood to protect it from direct sunlight and disturbance.
Ten choice-test arenas were placed in the vineyard for each of six trials, which were conducted on different dates. During each trial, arenas were placed at the base of vines near ant nests, separated from one another by at least 80 feet within a vine row and 24 feet between vine rows to ensure that only one colony fed from each. Nests were exposed to baits only once during the experiment. Ants were allowed to forage freely in the arenas for 24 hours. The arenas were then collected, and the average percentage of each bait removed was calculated. The percentage of a test bait removed from a single arena was calculated as the number of grams of a single bait removed divided by the total number of grams of all baits removed. Data were arcsine squareroot transformed and analyzed using analysis of variance and least significant difference to separate means within SAS.
On each of the test dates, the anchovy baits were removed from the test arenas in significantly greater amounts than all other baits (fig. 1). The average percentage removed of the two baits with anchovy plus imidacloprid varied slightly between test dates, but the differences were not consistent across test dates. This indicates that imidacloprid at the concentrations tested did not deter collection by the ants. During this experiment, field ant was observed foraging on Maxforce but at a very low rate.
Efficacy trial
We wanted to measure the short- and long-term impact of the most effective treatments for controlling field ant populations, as indicated by later foraging activity. In the efficacy trial, we tested both concentrations of imidacloprid (0.005% and 0.05%) in anchovy, Maxforce and a nontreated control. The experimental design was a completely randomized block, with four treatments replicated five times. Plots equaled about 0.04 acres and each consisted of 16 vines. In each plot, each of the 16 vines was treated with either concentration of imidadoprid in 20 grams of anchovy or 1 gram of Maxforce in a 90 ml. plastic vial placed and left at the base of the vine.
Pre- and post-treatment foraging activity of field ant were monitored using 20 grams of blank anchovy bait (no toxicant) in 90 ml. plastic vials placed at the base of the four central vines within each plot. After 24 hours, the vials were collected and returned to the laboratory, and the average amount of bait removed was calculated. Monitoring was conducted 2 days before treatment, and 10, 18, 93 and 122 days after treatment (DAT).
At 10 DAT, foraging activity of field ant significantly decreased in plots treated with Maxforce and anchovy plus 0.05% imidacloprid, but the foraging activity returned to the pre-treatment level by 18 DAT (fig. 2). Maxforce significantly reduced foraging activity below that of the control at 93 and 122 DAT. At 122 DAT, both anchovy baits reduced the foraging activity similarly, however only the lower rate of imidacloprid (0.005%) reduced the foraging activity significantly lower than the control.
Foraging activity
The process by which ants share food may explain these results. Larval stages, especially the later instars, are the only members of a colony that can feed on solid foods. Foragers bring solid food into the nest and place it on the mouthparts of the larva; the food is then chewed and digested. The larval stage uses much of the nutritional value for growth, but excess soluble proteins and amino acids can be stored (often in the salivary glands) and then later secreted to workers as a liquid food (Abbot 1978).
The decrease in foraging activity observed at 10 DAT suggests that the workers that foraged and processed the Maxforce or anchovy plus 0.05% imidacloprid baits suffered significant mortality; the rebound occurred as a result of the dead workers being replaced by workers from within the nest. The more delayed reduction in foraging activity observed at 93 DAT (Maxforce) and 122 DAT (Maxforce and anchovy plus 0.005% imidacloprid) strongly suggests that these baits affected significant mortality among the brood and reduced the colony size.
Developing bait stations
We are encouraged by these results. In subsequent experiments, we will vary both the percentage of imidacloprid in the anchovy bait and the application rate to determine if we can improve its efficacy. The advantage of formulating anchovy bait with imidacloprid is that imidacloprid is currently registered for use on grape as both a foliar and systemic insecticide. We believe that a bait such as this would quickly be approved for use against field ant as a broadcast bait, thereby eliminating the need for bait stations. The reduction in foraging activity in the plots treated with Maxforce suggests that hydramethylnon is also an effective toxicant. Experiments conducted in the spring and summer 2004 included various rates of hydramethylnon formulated in the anchovy bait.
An advantage to the commercial ant baits is their availability once registration is complete. Our data indicates that Maxforce with 1.0% hydramethylnon is also a candidate for such registration, although we have not yet developed a cost-effective bait station to deliver it. Densities of field ant can be high, with approximately 50 nests per acre. Our goal is to develop a disposable bait station that can be filled with an appropriate bait, such as anchovy or Maxforce, and placed near the entrance of each nest.
At this stage in our research, we do not fully understand how populations of other ant species — such as acrobat ant and the two species of fire ants — will respond in the absence of field ant. At no time during the preference trial did we observe acrobat ant foraging in the test arenas. However, the fire ants (S. molesta and S. xyloni) foraged Combat, and S. molesta also collected Niban. These baits may also be candidates for registration in bait stations if the population of any of these species were to increase to pest status in the absence of field ant.
Argentine ant baits and efficacy
In previous research, Rust et al. (2000) designed the choice-test arena described above in order to determine food preferences of Argentine ant. They found that both 20% sucrose water and Maxforce granular (nontoxic blank) were highly preferred and consistently collected by Argentine ant year-round. Rust et al. (2000) also determined the effective concentration range of several toxicants in sucrose water for Argentine ant. On the basis of their research, we chose a liquid bait (25% sucrose water plus 0.0001% thiamethoxam) and Maxforce to test in citrus.
The baits were tested in a 40-acre Valencia orange orchard. The liquid bait, liquid-plus-solid baits and controls (nontreated) were randomly assigned to twelve 1.5-acre plots such that each treatment was replicated four times. Each plot consisted of a 9-by-12 block of trees. Plots were separated by seven rows of trees, which served as a buffer. Baits were placed in stations at the base of every fourth tree in a plot along the irrigation line, providing an equivalent of 22 stations per acre.

Ant bait stations submitted to the U.S. Environmental Protection Agency for use in citrus orchards and grape vineyards were: (A) liquid, constructed from PVC pipe — design used in trials to control Argentine ant on citrus; (B) granular (Kness Manufacturing Co., Albia, IA); (C) granular, B&G Perimeter Patrol System (B&G Equipment Co., Plumsteadville, PA); (D) liquid, used to control Argentine ant in citrus (Whitmire Micro-Gen, St. Louis, MO); and (E) liquid, (Km AntPro, Nokomis, FL). All except (A) are commercially available.
Ant activity was estimated in each plot twice monthly using 50 ml. monitoring tubes constructed according to specifications described by Klotz et al. (2003). Tubes were filled with 25% sucrose water, weighed and taped to the trunk of each of nine trees located in a three-by-three configuration at the center of each plot. Two additional control tubes were hung in two of the central trees to measure evaporation. To prevent ants from foraging on the control tubes, they were suspended from string coated with Stickem Special (Seabright, Emeryville, Calif.). Ants were allowed to feed from the monitoring tubes for 24 hours. The monitoring and control tubes were then collected and reweighed to calculate weight loss. The evaporative water loss from control tubes was determined and used to calculate the net consumption from each monitoring tube, which provided us with an estimate of the activity of ants in each plot. During the 24-hour monitoring period, the toxic bait stations were temporarily sealed with plastic bags to prevent competition with the monitoring tubes.
Data was pooled across observation dates and analyzed using the Kruskal-Wallis nonparametric procedure (Systat version 9) to test for a treatment effect, and the Nemenyi procedure was used to test for differences among the treatments. During the pre-treatment observation period (May 14 through July 1), consumption of sucrose water did not differ significantly among the experimental plots (P = 0.99925). Post-treatment consumption of sucrose water, however, did differ among the plots (P < 0.0001). The plots treated with the liquid and liquid-plus-solid baits had less consumption of sugar water than the control, but did not significantly differ from one another (fig. 3).
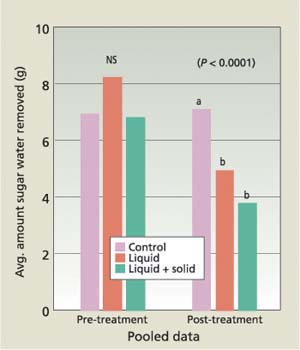
Fig. 3. Mean grams of sucrose water removed from monitoring tubes. Data was pooled across observation dates. Monitoring tubes were placed on citrus tree trunks and Argentine ants were allowed to feed for 24 hours. Means with the same letter are not significantly different.
Registration considerations
All pesticide formulations must be registered on a crop before they can be used legally to control a pest species, an essential consideration for taking control technologies from research and demonstration projects to production agriculture. To achieve registration, the agricultural chemical companies that handle the active ingredients must first agree to their use; their pesticides cannot be used without consent. Finally, the U.S. Environmental Protection Agency (EPA) must approve the registration, which normally requires data to be submitted. These data requirements can be extensive, and agricultural chemical companies normally bear the expense.
The Interregional Research Project Number 4 (IR-4) was organized to facilitate pesticide registration in cases where economic incentives for the companies are lacking. Such registrations have been termed “minor use.” For minor use, the costs of achieving registrations are normally more than the potential economic benefit to the companies. Ant baits for use in vineyards and orchards would be considered by most of the companies to be minor uses.
In cooperation with University of Hawaii and UC Riverside researchers, IR-4 has been working on ant baits in pineapple fields, vineyards and orchards. IR-4 negotiated with the EPA for concessions that will make it easier to register ant baits. In lieu of a broadcast application, ant baits will have to be delivered in bait stations placed in the pineapple field, orchard or vineyard, and the bait stations must prevent any potential contact of the pesticide with the harvestable crop. The degree of protection provided by the bait stations convinced EPA that certain expensive data requirements could be waived, making future registration of these innovative technologies much more likely.
Effective ant control
The management of ants that tend homopteran pests is a key component of integrated pest management in vineyards and citrus orchards. Low-toxicity ant baits are more effective than broad-spectrum insecticides because a bait is shared among nest mates and the queen(s). Additionally, ant baits are target-specific and when applied in stations, the risk to nontarget organisms and risk of environmental contamination is minimized.
Although we focused our study on developing baits to control ant pests in vineyards and citrus orchards, the methods we have developed to screen potential baits for feeding preference and test their efficacy under field conditions are applicable to other cropping systems. Our goal is to provide growers with cost-effective, easy-to-use bait delivery systems, and through collaboration with IR-4 to obtain registration of these products for use in agriculture.
Low-toxicity baits control ants in citrus orchards and grape vineyards

