


All Issues
Success of mite-fighting tactics evaluated: In Central Valley, early-season release of Willamette mites confers resistance on wine grapes
Publication Information
California Agriculture 52(6):21-24. https://doi.org/10.3733/ca.v052n06p21
Published November 01, 1998
PDF | Citation | Permissions
Abstract
Wine grapes in the Central Valley serve as hosts for several species of mites. Because these species have radically different economic effects, their correct identification is essential. Field trials revealed that some management practices were far more successful than others. For instance, treatments with dicofol (a miticide) and car-baryl (for leafhoppers) produced resurgences of mite populations to damaging levels. Releases of predaceous mites provided inconsistent control of herbivorous mites, and cover crops did not provide any clear advantages for mite management, although they may have other benefits. However, releases of Willamette mites at low densities early in the growing season consistently reduced chronically high populations of Pacific mites throughout the season. (Willamette mites, though they can become pests, are not damaging at low densities.) Scientists do not fully understand how Willamette mite releases confer plant protection, but evidence suggests they stimulate the host plant to reduce fecundity and survival of Pacific mites. Manipulations of host plant resistance offer new possibilities for control of pests such as spider mites.
Full text
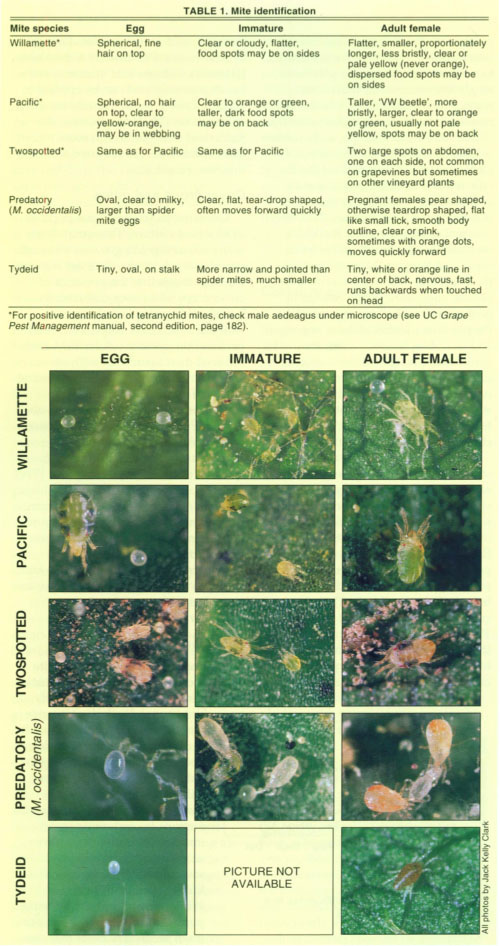
Central Valley wine grapes commonly support two species of Tetranychid spider mites, the Pacific mite and the Willamette mite (Flaherty et al. 1992). These are both herbivores that feed on leaf cells. In addition, grapevines support smaller tydeid mites, which eat pollen and fungal spores, as well as predatory mites, which consume both spider mites and tydeids.

A zinfandel vine that has been defoliated by high populations of Pacific spider mites near Lodi.
Correct identification of the mites is important because the species have different economic effects. Tydeids and predators are beneficial. Both Pacific (Tetranychus pacificus) and Willamette (Eotetranychus willamettei) mites have the potential to become pests, although grapevines can tolerate considerable spider mite feeding by either species before treatment is economically justified. Pacific mites are more damaging at lower densities than Willamette mites (Flaherty and Huffaker 1970; Kinn and Doutt 1972). However, even Pacific mites must exceed 8 to 14 mites per leaf (or 50% to 65% infested leaves), on average, before a treatment may be needed (Flaherty et al. 1992). Willamette mites must exceed 30 mites per leaf, on average, before any economic losses have been detected (Welter et al. 1989). Before treating mites, predator density, host plant related mortality, and other sources of mortality must be considered, as these can reduce mite populations without chemical intervention before economic damage is achieved.
Pacific mites at damaging densities cause the leaves to appear bronzed, especially between leaf veins, and can distort shoot tips. If they are permitted to build up to very high densities, they may defoliate the vines. Pacific mites slow the rate at which the berries sweeten, forcing growers to either harvest at a suboptimal sugar content or leave the crop on the vine into the fall when seasonal risks of spoilage increase. Varieties such as zinfandel grapes that express less vigorous vegetative growth appear to be particularly susceptible to damage by spider mites. Pacific mites tend to cause problems where vines are water stressed or where disruptive pesticides such as dicofol (for spider mites) or carbaryl (for leafhoppers) have been used. Densities vary from year to year, perhaps occurring at lower densities when summers are relatively cool with abundant moisture (Karban et al. 1997). Pacific mites tend not to build to populations where they become economic problems in vineyards that have populations of Willamette mites or populations of predatory mites, especially when the Willamette mites or predators are present early in the season.
Willamette mites are ubiquitous in vineyards and in natural riparian corridors that contain native grapes (Karban et al. 1997). Willamette mites — unlike Pacific and twospotted spider mites — have a narrow host range that includes cultivated and wild grapes and relatively few other species. As a result, Willamette mites spend their entire life cycles associated with grapevines.
At high densities, Willamette mites can cause yellowing between the leaf veins on light cultivars and reddish discoloration on dark cultivars, but generally not the reddish bronzing associated with Pacific mites. In certain situations — such as after a disruptive pesticide spray — their populations can build to economically damaging levels. Some vineyards in cooler, wetter areas, such as the Sierra foothills and some areas of the southern San Joaquin Valley with sandier soils, experience damaging populations of Willamette mites more frequently. Willamette mites will cause no significant reductions in berry sugar concentration when populations are below 30 mites per leaf, and 50 mites per leaf may be a more accurate threshold on more vigorous cultivars (Welter et al. 1989). Like Pacific mites, high densities of Willamette mites often follow dicofol sprays for spider mites and disruptive insecticides applied for leafhopper control.
Chemical controls
Chemical sprays remain the most frequently used tactic to control mites. Two acaricides — dicofol and propargite — are commonly used in California for grapes. We have conducted experiments with these two miticides in vineyards containing high densities of Pacific mites. Dicofol was found to reduce mite numbers immediately following its application. This effect and the fact that current law does not require a lengthy period after application before workers may re-enter the field has made it appealing to growers. However, these advantages are more than offset by the consistent and strong resurgences in mite numbers that follow dicofol applications (fig. 1) (Karban et al. 1997). In the vineyard depicted in figure 1, reductions in sugar content were associated with the mite resurgences (fig. 2). The reason for resurgences is unclear; dicofol may kill predatory mites, causing herbivore populations to explode, make plants more susceptible or stimulate the mites themselves to increase.
Although current law requires a period of 28 days after propargite use before workers may re-enter treated vineyards, more growers are trying the newer chemical because it has not been reported to produce mite resurgences. Resurgences of mites have been observed after the use of carbaryl for leafhopper control (fig. 3). As a result, dicofol should not be used, and particularly disruptive insecticides such as carbaryl should be avoided in vineyards where there is a risk of mite problems.
Predatory mites
Predatory mites consume both Pacific and Willamette mites and can provide biological control of these herbivores. The most commonly found predatory mite species in the Central Valley is the western orchard predator mite, Galendromus (=Metaseiulus) occidentalis, although other native species such as M. mcgregori can be important locally. G. occidentalis can be purchased from commercial insectaries and farm supply companies to augment naturally occurring populations. In numerous experiments, we have released G. occidentalis into vineyards with high populations of herbivorous mites. The predatory mites provided inconsistent control of Pacific and Willamette mites (English-Loeb et al. 1993; Hanna et al. 1997). The most cost-effective biological control of spider mites in vineyards is to promote the development of naturally occurring predator populations by preserving alternate prey such as tydeid mites (Flaherty and Hoy 1971) and by using only selective pesticides when necessary.

Fig. 1. Densities (mean ± 1 s.e.) of Pacific and Willamette mites in a zinfandel vineyard in 1992, one year after application of dicofol for mite control.
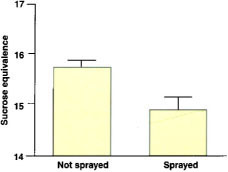
Fig. 2. Berry sugar concentrations (mean ± 1 s.e.) in 1992 associated with the treatments shown in fig. 1.
Willamette mite releases
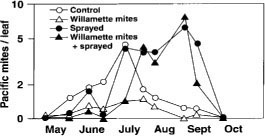
Vineyards that have chronically high populations of Pacific mites generally also have very low populations of Willamette mites and predatory mites. We have released Willamette mites into such vineyards early in the season and thereby lowered densities of all mites with better effect than other control options (fig. 4) (Karban et al. 1997). We do not understand which physiological or biochemical changes in the grapevines are responsible for induced resistance against Pacific mites, although survival and especially fecundity of the mites are reduced. In laboratory trials, fecundity of Pacific mites was reduced by 65% on leaves of plants with induced resistance, and survival of adult females was reduced by 56% relative to uninduced controls (Karban, unpublished data).
Although this tactic was consistently effective, there are problems with implementation because Willamette mites are not commercially available and are difficult to distinguish from Pacific mites and other related species. Therefore we conducted experiments to determine whether a single introduction of Willamette mites was sustainable or whether they needed to be reintroduced repeatedly.
We found that a single, early season release of Willamette mites was sufficient to establish and replace the Pacific mites during the entire season, and even into subsequent seasons if not disrupted by pesticides. This effect was just as strong even if the Willamette mites and Pacific mites were separated temporally or spatially on the plant and even if predators were absent, suggesting that a change in host plant quality was responsible. Proper mite identification is important and sampling for both mites should be continued throughout the season to determine if either population has reached a treatment threshold. Early season releases should not be attempted in vineyards that experience chronic Willamette mite pressure.
Feeding by Willamette mites as the shoots were expanding early in the season reduced the quality of the foliage for mites that fed later in the season. Damage by Pacific mites at budbreak also induces resistance (English-Loeb et al. 1998). Recent experimental results in other systems suggest that jasmonic acid is the plant hormone responsible for these increases in plant resistance (Thaler et al. 1996). This role for jasmonic acid as the signal for damage by herbivores is highly conserved in most plant species. All of the plants we examined use jasmonic acid as a signal for damage by herbivores. Preliminary experimental results indicate that minute quantities of jasmonic acid can be applied to grape leaves to induce resistance to spider mites (Thaler and Omer, unpublished data), although more rigorous experiments are required to assess this new possibility.

Fig. 3. Mean densities of grape leafhoppers and Willamette mites following an application of carbaryl for leafhopper control in 1995 in a merlot vineyard near Lodi.
Cover crops and mites
Cover crops have become popular in vineyard culture. These may have many advantages to growers although preliminary evidence does not support the hypothesis that the presence of cover crops will prove consistently beneficial for mite management (Karban et al. 1997). Improved water penetration, greater soil fertility, and reduced dust associated with cover crops may be responsible for reducing effects of spider mites in those instances where benefits have been observed. Further work and experience will be required before a more informed assessment of the consequences of cover crops for spider mites in vineyards is possible.
Managing mites does not occur independently of other vineyard, horticultural and pest management practices. Proper irrigation, nutrient management, and dust control in and around vineyards are essential to manage mites as is careful use of non-disruptive pesticides.
Mite identification key
Distinguishing between the mite species found in vineyards is essential to managing them effectively. Unfortunately, these mite species are very small and difficult to distinguish, yet proper detection and identification is essential for effective management (table 1). The UC Grape Pest Management manual contains directions for mounting male mites for unambiguous microscopic identification as well as several photographs of selected life stages. Pacific and Willamette mites both have the potential to be pests. Mites are often secondary pests that can be avoided by alleviating water stress and dust in vineyards, or substituting other control tactics for disruptive miticides such as dicofol and insecticides such as carbaryl. In many of the vineyards in which we have worked over the past 10 years, mite problems were greatly diminished after a year of no application of miticides. Grape growers with chronic devastating problems caused by Pacific mites may wish to try introducing low densities of Willamette mites soon after budbreak, avoiding subsequent applications of miticides.
Success of mite-fighting tactics evaluated: In Central Valley, early-season release of Willamette mites confers resistance on wine grapes

