


All Issues
Cutting off irrigation early may reduce almond hull rot
Publication Information
California Agriculture 48(3):33-36.
Published May 01, 1994
PDF | Citation | Permissions
Abstract
Cutting off irrigation two weeks before harvest substantially reduced the damage caused by hull rot in a Kern County almond orchard trial. The extent of leaf death was affected by the stage of hull split at which the fruit were inoculated, but not by the concentration of inoculum.
Full text

Above, the incidence of almond hull rot was affected by the timing of irrigation cut-off and hull split size, but not by inoculum concentration. The incidence of hull rot was significantly higher in trees irrigated within 2 weeks of harvest (the greener trees shown above). Right, almond yields are reduced by hull rot, which destroys the fruiting wood.
Hull rot is a sporadic but sometimes serious disease of almond, Prunus dulcis (Mill) D. Webb, in California. It is caused by Rhizopus stolonifer (Ehrenb:Fr.) Vuill. or Monilinia fructicola (G. Wint.) Honey. Three other fungi, R. circinans Tiegh, R. arrhizus A. Fischer and M. laxa (Kaderhold & Ruhland) Honey, can cause hull rot symptoms, but rarely are found associated with the disease in the field. Although all almond cultivars are susceptible, the widely planted ‘Nonpareil’ cultivar is the most susceptible. Vigorous, heavily cropped orchards usually sustain the greatest damage, and no control measures have been defined.
The first symptom of hull rot is a grayish lesion, which soon turns tan to brown on the hull of the maturing almond fruit. Then the nearby leaves and part or all of the spur or shoot upon which the infected fruit is borne begins to die, probably due to a toxin produced by the pathogens and transported from the infected hull to the spurs and leaves. The vascular tissues leading to the infected fruit die and turn black. The dead leaves remain attached to the twigs, imparting a scorched appearance to badly damaged trees. Sporulation by Rhizopus is easily visible between the hull and shell, and M. fructicola often can be found sporulating on inner and outer hull surfaces. Although the edible nut is not harmed, yields are reduced because hull rot destroys the fruiting wood.
Since pathogens cannot invade the outer hull surface of the almond fruit, hull rot infections occur during hull split, when the hull opens along the suture. Infection starts as the fungal spores, probably carried by insects and wind currents, are deposited on the inner hull surface. The period of potential infection corresponds to the hull split process: Hull split begins in early July in ‘Nonpareil’ trees planted in the southern San Joaquin Valley; most nuts on those trees have split hulls by late July, and essentially all hulls have opened by harvest, in mid-August. As the split enlarges over several days, the hull detaches from the pedicel and loses moisture. At harvest, the hulls of most fruit are fully open, dry and leathery. Healthy nuts remain attached to the tree by only a few vascular elements and are easily dislodged when the tree is shaken. Infected fruit cling to the tree after harvest and must be removed, at considerable expense, because they can serve as overwintering sites for navel orangeworm, Amyelois transitella (Walker), a serious insect pest of almond.

Hull rot infection begins as the fungal spores are deposited on the inner hull surface during hull split. The first symptom of hull rot is a grayish lesion, which turns tan to brown on the hull of the maturing almond fruit.
Hull rot is most prevalent in high-yielding, densely canopied orchards that receive ample water and nitrogen. Sudden outbreaks often follow late preharvest irrigations, which suggested to us that changes in irrigation practices might be useful in minimizing losses to hull rot. Our trials investigated the effects of irrigation cut-off date, hull split and inoculum concentration on the incidence and severity of hull rot.
Kern County experiments
We conducted the experiments in a commercial almond orchard in Kern County, California. The orchard had been planted in 1981 with ‘Nonpareil’ and ‘Carmel’ cultivars in a 2:2 pattern on a 25-feet-by-25-feet spacing. In 1989 an irrigation experiment began. In eight irrigation cut-off treatments, irrigation was terminated at weekly intervals beginning in late June and ending just before harvest in mid-August. The treatments were applied to the same trees for three consecutive years. Apart from irrigation, all orchard practices were standard for the region and maintained by the grower.
The hull rot experiments were conducted during two years — 1990 and 1991 — of the irrigation cut-off experiment. Before the onset of the irrigation cut-off treatments each year, all trees received full irrigation, estimated with the California Irrigation Management Information System (CIMIS) modified Pinman reference crop water use and almond crop coefficients. Water was applied by microsprinklers. The cumulative inches of preharvest water delivered to treatments one through eight were, respectively, 17.5, 19.3, 21.1, 22.9, 25.6, 27.4, 29.2 and 31.0 The intervals between the last irrigation and harvest were 52, 46, 39, 32, 25, 18, 11 and 4 days, respectively. The last irrigation in commercial orchards usually occurs within two weeks of harvest, a timing represented by treatments seven and eight.
Predawn leaf water potential readings were made on four trees in one replication of each treatment on July 26, and August 2, 9, and 16, 1991. Readings were made for one mature leaf per tree by excising and immediately pressurizing it in a portable pressure chamber.
We measured percent of hull split and moisture content to see whether either of these factors influenced development of the disease. Measurements were derived from 20 fruit selected at random from the periphery of each group of eight data trees. Four collections were made at approximately weekly intervals beginning at early hull split in July each year. Fruit were sealed in plastic Ziploc bags and stored on ice for return to the laboratory. Hulls were considered split if any part of the suture had separated 0.04 inch or more. Hulls were separated from the nuts, weighed, then airdried in a forced-air oven at 150°F for 72 hours to determine dry weight. Percentage hull moisture content was calculated from these figures.
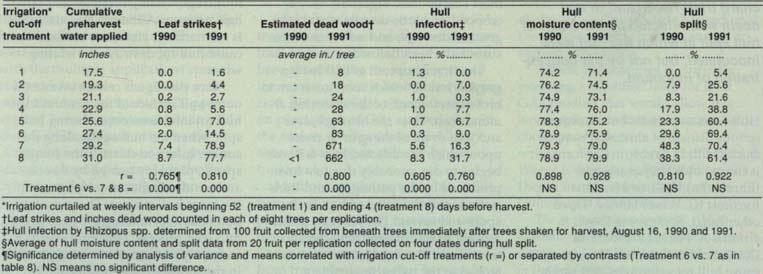
TABLE 1. Effect of irrigation cut-off on hull rot caused by Rhizopus sp., hull moisture content and hull split, Kern County, 1990–1991
There were three replications of each irrigation cut-off treatment, arranged in a randomized complete block design, and each plot consisted of eight rows 12 trees long. Data were collected from the center four pairs of ‘Nonpareil’ trees within each plot.
Hull rot was studied in two ways, by measuring natural infection and conducting inoculation experiments.
Natural infection
We measured the incidence of natural infection at harvest in 1990 and 1991. Trees were shaken on August 16 in both years. Immediately afterwards, strikes were counted (strike designated a short spur with one or two clusters of dead leaves or a single cluster of dead leaves on a shoot), and the inches of dead wood present were estimated in all eight data trees in each replication. Random samples of fruit were collected from the ground beneath the trees, placed in paper bags, and transported to the laboratory, where 100 fruit were drawn from each sample and examined for presence of hull lesions and growth of hull rot fungi.
Hull split size and inoculum concentration
We studied the effects of hull split size and inoculum concentration on incidence of hull rot in 1991. Inoculum was prepared from cultures of one isolate of R. stolonifer obtained from almond. Approximately 0.1 to 0.2 ml of inoculum was introduced into the fruit through open sutures with an artist's airbrush. No water-soaking or other damage to the hull tissues was observed. We inoculated 25 fruit per replication at approximately weekly intervals on each of four dates, beginning at early hull split and ending within 2 weeks of harvest. The inoculated fruit were numbered and their locations identified with flagging tape. Disease data were collected during the 5 days before harvest. The condition of leaves near each inoculated fruit was tabulated as healthy, missing or stricken, then the fruit were removed, returned to the laboratory and examined for hull infections and fungal growth.
The influence of hull split size on infection was examined in treatments six, seven and eight. Fruit with small (0.04–0.20 inch), medium (0.20–0.50 inch) and large (> 0.50 inch) hull splits, measured at the widest point of separation, were inoculated with 104 spores per ml suspension of R. stolonifer on the second, third and fourth inoculation dates. The first inoculation date was excluded because there were no fruit with large hull splits. Percent of hull moisture content was measured in 10 fruit from each of the three hull split sizes from each replication on each inoculation date.
Three levels of inoculum concentration were tested in 1991 for their effects on hull infection and leaf death. Fruit that had small- to medium-sized hull splits, in treatments four through eight, were inoculated with 103, 104 or 105 spores per ml suspensions of R. stolonifer. One group of 25 fruit with medium split hulls in each replication served as a noninoculated control for this experiment and the hull split size experiment described above.
Weather data were obtained from a CIMIS station located approximately 2.5 miles from the orchard.
Hull rot and irrigation
Hull rot incidence was affected by irrigation cut-off and hull split size, but not by inoculum concentration.
Natural infection
Hull rot caused by natural infection was reduced in early irrigation cut-off treatments (one through six) and was most severe in later treatments (seven and eight), in which one or two irrigations were made during the last two weeks before harvest (table 1). This distinct delineation between the earliest six and last two treatments was noted in both years for percentage hulls infected and number of strikes per tree, and also for estimated inches of dead wood per tree in 1991. However, neither percentage hull moisture content nor percentage split responded consistently.

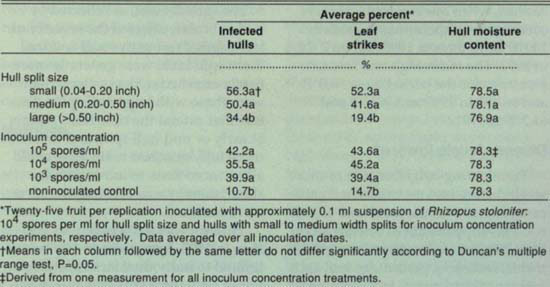
TABLE 3. Effect of hull split size and inoculum concentration on hull rot caused by Rhizopus spp., Kern County, 1991
Predawn leaf water potentials fell below -20 bars within 11 days of water deprivation in early treatments (table 2). A week before harvest, trees in later treatments were less stressed – registering values of -6 and -9 bars – than those measured in earlier treatments. Fully irrigated almond trees have predawn leaf water potentials of -4 to -7 bars. Only trees in the latest irrigation cut-off treatment (treatment eight) maintained low predawn leaf water potentials through harvest.
Hull split size and inoculum concentration
Fruit with large hull splits had fewer infected hulls and strikes, but not lower hull moisture content, than did fruit with small or medium hull splits (table 3). Inoculum concentration, excluding the noninoculated control treatment, did not affect the percentage of infected hulls or strikes (table 3).
Average daily maximum and minimum summer temperatures were higher in 1990 than in 1991. Through the course of the irrigation cut-off experiment, mid-June through mid-August, there were 56 and 38 days in 1990 and 1991, respectively, when temperatures rose above 90°F, and 16 and 6 days, respectively, when they rose above 100°F. The high and low means for that period were 97.7°F and 63.5°F in 1990 and 91.4°F and 60.4°F in 1991. During the five weeks before harvest, when our experiments were conducted, temperatures exceeded 100°F on 12 days in 1990 and on 2 days in 1991. The mean high and low temperatures for the period were 98.8°F and 66.4°F in 1990 and 92.7°F and 61.2°F in 1991.
Disease levels lowered
Terminating irrigation two or more weeks before harvest caused a significant reduction in the incidence of hull rot. This suppression of disease might have been related to any of several elements, including percentage hull split and moisture content, inoculum production and spread, or toxin production and transport. Differences in percentage hull split and hull moisture content did not account for the reduced levels of hull rot. The low percentages of split hulls found in irrigation cut-off treatments one through three may have contributed to the low levels of disease found in those, but in treatments four through eight hull split surpassed 70% three weeks before harvest. Thus most fruit were susceptible to infection in four, five and six, as well as seven and eight, yet only treatments seven and eight exhibited severe symptoms. Although hull moisture content may prove to influence the growth of R. stolonifer or the production of toxin, the small changes encountered among our treatments did not affect hull rot incidence.
The production and dissemination of inoculum might have varied among the treatments and consequently affected disease levels. There were obvious differences in tree condition among the treatments. Early treatment plots were dry, dusty and hot, and the trees showed the usual water stress symptoms consistent with the magnitude of the predawn leaf water potentials. These conditions may have been detrimental to the growth and sporulation of Rhizopus spp. Trees in the latest treatments were most lush, and their dense canopies provided more shade and a cooler environment. In addition, there was more weed growth under these trees. Insects are suspected vectors of the hull rot fungi, and climate might affect their biology and movement to the trees.
Hull attachment, as reflected by hull split size, affected the severity of leaf death. Fruit with small and medium split hulls were generally more firmly attached to the pedicels than were those with large hull splits. Factors that extend the time hulls remain at early or mid hull split, and therefore more fully attached to the tree, could allow more toxin to move into the shoot, thereby exacerbating the severity of hull rot.
Hull rot was more severe in 1991 than in 1990. Symptoms in 1990 were limited to individual strikes, and we found only three strikes in our inoculation experiments, all located in treatment eight. Collapsed shoots and small limbs as well as individual strikes were present in 1991. In both years most symptoms developed within the last two weeks before harvest. The high ambient air temperatures in 1990 (56 days with mean temperatures above 90°F) perhaps intensified water stress, and this may have affected hull split, the abscission process, and the transport of toxin. The pathogen, too, may have been limited by the hot weather. The reported maximum growth temperature for R. stolonifer is 95°F.
Manipulation of irrigation could be an important management tool for the control of hull rot. However, our results in Kern County may not apply to other orchard situations. The experimental orchard was planted on shallow soil with a relatively low water-holding capacity so water stress quickly followed water deprivation. Hull rot may not be as easily managed by cessation of irrigation in orchards on deeper soils, because trees will be able to obtain water from lower soil depths in the absence of irrigation. Less stringent deficit irrigation schemes that limit water over the irrigation season rather than summarily cut it off also may not be effective. These situations are the subjects of further investigation.
Cutting off irrigation early may reduce almond hull rot

