


All Issues
Researchers take a look at the versatile L.: wanton weed and/or golf turf?
Publication Information
California Agriculture 46(3):24-26.
Published May 01, 1992
PDF | Citation | Permissions
Abstract
Poa annua L. (annual bluegrass) is seen by researchers as both a troublesome' weed, especially on golf courses, and as an attractive turf. A wealth of genetic variations in the morphological, physiological and life history characters of this weedy species has been detected. Awareness of the genetic differences in growth habit and reproductive strategies between annual and perennial biotypes of P. annua can be helpful in managing it as turf, as well as in situations where it is treated as a weed.
Full text

On left, annual bluegrass from golf course rough displays an upright growth habit characterized by few tiller branches and more flowers. On right, bluegrass from golf green displays prostrate growth habit with profuse tiller branches.
Research conducted on Poa annua L. (annual bluegrass) in the United States has either focused on it as a weed requiring effective control or on its adaptation as a turfgrass. As a weed, it is widespread in California turf and landscapes, especially on golf courses. On the other hand, perennial, prostrate-growing P. annua strains have been suggested for use on golf greens, despite the fact that P. annua turf has low resistance to drought and heat, and frequently deteriorates during dry, warm summer months.
Attempts to select for heat resistance in P. annua reveal no correlation between heat resistance and geographic distribution. It is possible, however, that plants complete their life cycle before the onset of heat stress and that heat resistance is therefore associated more with management practices than with climate. Research into management practices that may affect control and/or performance of this species seems, therefore, reasonable.
Regardless of how much P. annua is improved by breeding and selection programs, its characteristic variability will ensure its continuance as a weedy grass problem. This, however, should not discourage future research. Without knowledge of how the variable genetic characters and turf management practices interact, it is not possible to establish effective control programs or a breeding program for this species.
Seed dormancy and life history
Golf courses are particularly convenient sites for studying P. annua, because they are home to many turf management schemes. Distinct P. annua biotypes can readily be identified under different turf management practices. The annual biotype is often found in drier, less intensively managed areas; the perennial biotype commonly thrives in wet, highly managed areas. The two major management practices on a golf course, irrigation and mowing, provide a gradient of intensity of turf management practices. Higher management means more frequent mowing and irrigation as well as closer mowing set. Poa annua populations from the Davis Municipal Golf Course in Davis, California were examined to detect the effects of management practices and physical environment on seed dormancy and seed banks, and on genetic differentiation and adaptation of life history traits. Also studied were the effects of light intensity and growth regulators on P. annua biotypes.
Seed dormancy and seed bank.
Seeds of P. annua were collected from areas of three turf management categories: golf green, fairway and rough. Seeds were tested for the effects of temperature on seed germination and seed bank of P. annua. Seed germination was examined at 68° and 52°F under 24 hours of light in growth chambers.
After a 20-day incubation, highly significant differences in seed germination rate appeared between exposures to high and low temperatures and between green, fairway and rough. Between-site differences within a turf management category were not significant. A positive, significant correlation in seed germination between field-collected seed populations and their respective greenhouse-produced seed progenies was also observed (fig. 1).
At 68°F, seed germination in the rough, which receives no irrigation, was uniformly low, less than 20%. Conversely, germination in the golf greens, which receive frequent irrigation, was high, greater than 80%. A wide range of seed germination was found in seed families in fairways, which receive infrequent irrigation and mowing. Highly significant differences were found between turf management categories, and between families within a management category. Conversely, at 47°F, seed germination was uniformly high, and no difference could be distinguished between golf green and rough populations.
These studies indicate that the high temperature-enforced seed dormancy differences existing among golf course P. annua populations are associated with turf irrigation and mowing. Variation in seed germination abilities in relation to temperature has been reported among populations of P. annua collected from a wide range of geographical areas, suggesting that germination variations may be ecologically significant. It may be that temperature triggers seed germination and maximizes chances for survival.
In golf course roughs, P. annua is not able to survive the warm, dry, summer months of California’s Central Valley. Although seeds may occasionally receive water from runoff or drift, it will not be sufficient to support the plants’ life cycle, if germination does occur. The Central Valley’s seasonal precipitation changes have a regular and predictable pattern; rainfall is greatest in the winter and spring when the monthly mean temperature is lowest. Therefore, not surprisingly, seed germination of rough populations is controlled by temperature rather than by moisture.
Populations of P. annua on golf greens receive frequent irrigation, assuring seeds of sufficient moisture for their survival, regardless of when they germinate. Temperature-dependent seed germination would, therefore, be at a disadvantage on golf greens.
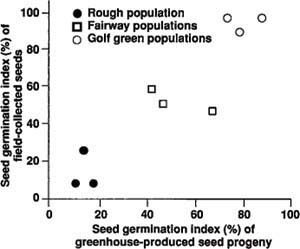
Fig. 1. The joint distribution and correlation of seed germination index between the field-collected Poa annua seed populations and their respective greenhouse poly-crossed seed progeny populations.
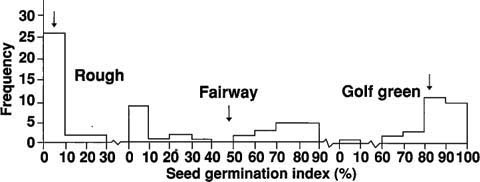
Fig. 2. The distribution of seed germination index in the three golf course Poa annua populations under different irrigation and mowing practices (means denoted by arrows).
The bimodal distribution of seed germination (fig. 2) found in fairways suggests that they are heterogeneous habitats rather than an intermediate uniform environment distinctly differing from golf greens and roughs. Evidence for this form of selection in P. annua has been reported by other researchers for plants growing on bowling greens subject to clipping and those in adjacent flower beds. Perhaps selection for temperature-enforced seed dormancy has been operating among the golf course annual bluegrass populations.
Poa annua life history characters. Plants collected from two sites, a golf green and a rough, were grown in similar but different greenhouses, to prevent cross-fertilization between biotypes by air currents. Each biotype (a group of plants) was allowed to produce seeds, and after further inbreeding each was grouped into families. Seeds were germinated, and seedlings were grown in a growth chamber at 61° F days and at 53°F nights with a 12-hour photoperiod at a light intensity range of 350 to 450 microeinsteins m-2 sec-1. In each family, half of the plants were harvested at flowering. The remainder were harvested at maturity, arbitrarily chosen as 60 days after the first spike appeared. Data were recorded at flowering and at maturity for the following characters: flowering time (number of days from germination to flowering), number of tillers (shoot with their roots) and inflorescences (the flower-bearing structures of grasses), number of nodes per flowering tiller, and vegetative and reproductive dry weights.
Green and rough populations of P. annua differed significantly for all traits except flowering time. The flowering and reproductive characteristics at maturity were greater in the rough population; the number of vegetative shoots was greater in the green population (table 1). Reproductive output was greater in the rough population, and plants had more flowering characteristics (number of tillers, number of nodes per tiller, number of inflorescences and dry weight). Comparisons of life history characters within families show that genetic variation is usually greater in the green than in the rough population, while between-family variation is greater in the rough than in the green. Total genetic variation is almost always greater in the rough than in the green.
In P. annua, young shoots develop at the nodes of the crown (unelongated stem) and at the nodes of prostrate tiller shoots. Nodes per shoot are distributed from the base to the tip of the shoot. As a result, younger shoots have fewer nodes. Number of nodes and tillering (developing new shoots) was greater in the rough than in the green population. Flowering was more abundant in the rough.
Most of the characters measured were highly heritable, indicating that a substantial portion of the variability observed between populations in life-history parameters of the P. annua is genetic in origin. The most probable explanation for the genetic differences recorded in this study is selection produced by turf management practices.
Poa annua from high adult mortality environments (rough) tend to have an annual life history; plants from high juvenile mortality environments (green) have a perennial life history. This suggests that the green (perennial) population could have evolved from the rough population. Such evolution would have had occurred relatively rapidly, as this golf course was approximately 25 years old. This suggests that for P. annua, selection forces of turf management act through both physiological and developmental constraints, and these constraints can channel evolution of this species to very localized biotypes.
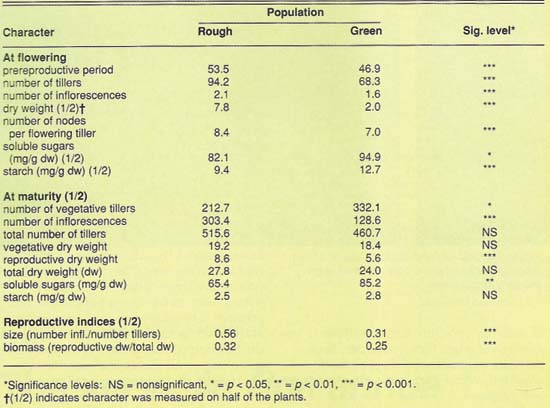

TABLE 2. Effect of light intensity and Paclobutrazol treatment on dry weight partitioning of Poa annua biotypes
Effect of light and growth regulator
The growth regulator Paclobutrazol is used in landscape management to restrict plant growth. Because in many situations P. annua tends to thrive more in shade than in sun, effects of both the growth regulator and shade on P. annua growth were investigated. Poa annua plants of both green and rough biotypes were grown in the greenhouse. Nine combinations of light intensity (full sun, one-half full sun, and one-third full sun) and the growth regulator (control, 3 mg or 90 mg per 90 cm2) treatment were used. The first application of Paclobutrazol was at the beginning of the experiment; the second application was 6 weeks later. Plants were harvested at the end of 12 weeks, and shoot, root and inflorescence dry weights were recorded.
Plants from golf greens produced greater total plant dry weight than rough biotypes under all light and growth regulator treatment combinations (table 2). Reducing light intensity from full to one-half full sunlight increased total dry weight for both green and rough biotypes. Paclobutrazol treatment reduced dry weight production considerably for both green and rough biotypes. With the treatment, both biotypes produced more dry weight under low light than under high light intensities. Lower light intensity and growth regulator treatments increased inflorescence dry weight partitioning of the rough biotype, but had no significant effect on the golf green biotype. Root growth was reduced, but dry weight partitioning was increased. Overall, root growth was less affected by the growth regulator than shoot growth for both biotypes.
Conclusions
There is a wealth of genetic variation in the morphological, physiological and life history characters of P. annua. Turfgrass management practices and environmental conditions act as selective forces and can maintain a distinct genetic difference between P. annua populations at the microecological level. Knowledge of the interaction between environment and seed germination in P. annua, such as the temperature-enforced seed dormancy difference, is important in the effective use of preemergence herbicides. In certain circumstances, P. annua has been adapted as a naturally invasive turfgrass. Awareness of genetic differences of growth habit and reproductive strategies (vegetative and sexual) between annual and perennial biotypes of P. annua at the microecological level can be helpful in establishing a management program in these situations, as well as in cases where the species is treated as a weed.
The growth regulator Paclobutrazol can effectively inhibit P. annua growth. However, it should be used with extreme caution, because it may severely damage existing turfgrass. More research is needed to discover differential response to this growth regulator among turfgrass species before it can be recommended for controlling P. annua in existing turf.
L. Wu is Professor, J. A. Harding is Professor and M. Borgonovo is Visiting Scientist from Italy, Department of Environmental Horticulture, UC Davis. M. A. Harivandi is Farm Advisor, Alameda County.
This research project was supported by research awards offered, by the Northern California Turfgrass Council, the Elvenia J. Slosson Endowment Fund and Agricultural Experiment Station Project, UC Davis, Ca-EHT4063-H.
Researchers take a look at the versatile L.: wanton weed and/or golf turf?

