


All Issues
Predatory mites help control thrips on floriculture crops
Publication Information
California Agriculture 44(6):19-21.
Published November 01, 1990
PDF | Citation | Permissions
Abstract
Western flower thrips is an increasingly important pest of field and greenhouse crops. Biological control with predaceous mites may be a viable option when dealing with this difficult pest.
Full text

Immature western flower thrips feed in the central, terminal growth point of this chrysanthemum, where they are protected from most pesticides.
The western flower thrips (WFT), Frankliniella occidentalis (Pergande), is one of the most serious worldwide pests of floricultural crops. Several factors associated with its biology and the mechanisms through which it damages flower crops have contributed to a sudden increase in its seriousness as a pest in the field and greenhouse.
WFT has a very large host range, a rapid development cycle, and a high reproductive rate. Many weeds and field crops serve as hosts for the pest, so migration into greenhouses is a continual problem. Because thrips are small and because they feed deep within young, terminal foliage and developing flowers, early detection is difficult. Visible damage from WFT is easily identified, but it often goes undiscovered until a large population is present in the greenhouse.
WFT's feeding position deep within the plant affords it considerable protection from insecticides. This and the development of insecticide resistance in many California greenhouse populations have reduced the feasibility of control using insecticides. Spraying open flowers is often complicated by problems with phytotoxicity and residue. Finally, WFT is one of the few known vectors of tomato spotted wilt virus, which affects many major floricultural crops (e.g., chrysanthemums, New Guinea impatiens, and gloxinia).
Although insecticides are difficult to use and present their own unique problems, they remain the grower's primary line of defense. One alternative that has not been explored in floriculture is the use of natural enemies for control of WFT. Biological control is generally more difficult in floriculture than in other segments of agriculture because of the crops' aesthetic value. However, successful examples can be cited for a number of different ornamental crops (see California Agriculture March-April 1984, January-February 1986, March-April 1988).
We initiated a study to evaluate the use of two predatory mite species, Amblyseius cucumeris Ouds. and A. barkeri (Hughes), for control of WFT on floricultural crops. These mites have successfully controlled onion thrips (Thrips tabaci Lind.) on glasshouse crops including sweet pepper in Europe, but little information is available on their potential for control of WFT on ornamental crops.
Introducing and culturing predatory mites
We obtained A. cucumeris and A. barkeri from three sources: the Danish Research Center for Plant Protection; Koppert BV. of the Netherlands; and Applied Bio-Nomics of British Columbia. For culturing, we used grain mites (Acarus farris [L.]) as a substitute prey for both predatory mites. We started a culture of the mite by adding some of the material containing only the prey to a medium of bran and wheat germ. Glass beakers were filled with this mixture and then placed into loosely covered glass jars containing a concentrated solution of potassium nitrate. This solution maintained a relative humidity (RH) of 90 to 95% necessary for grain mite reproduction. Two weeks later, when grain mite populations had reached sufficient levels, 50 individuals of a predator species were added to each culture beaker. Jars containing A. cucumeris or A. barkeri cultures were placed in separate environmental chambers at 21° to 22°C and 65 to 95% RH. An automatic humidifier maintained a high relative humidity in each chamber.
After about 20 days, the culturing vessels had built up large populations of A. cucumeris and A. barkeri. We aspirated adults off the sides of the containers and into small straws for field release.
Trial I
For the first trial, we grew chrysanthemum plants (cultivar ‘Hurricane’) from rooted cuttings and placed them into 15 cm diameter plastic pots in a greenhouse. After 3 weeks we selected 30 healthy plants of similar heights carrying 12 to 16 leaves each, and we trimmed each plant to only 10 mature leaves. Tanglefoot was applied at the base of each plant to prevent the thrips and mites from moving down the stem and into the soil. The plants were exposed to WFT for 7 days in a small glasshouse (9.7 m2) where the thrips were being cultured on roses.
After exposure, we transferred all plants to another, larger greenhouse (133 m2) with normal fluctuating temperature (6.7° to 35°C) and relative humidity (30 to 40%). Plants were placed in random groups of ten under separate muslin screen cages (each cage served as a treatment). The first cage was a predator-free control, and the second and third received A. cucumeris or A. barkeri, respectively. The day that the plants were transferred to muslin cages, we used a hand lens to count thrips on the upper and lower surfaces of all ten mature leaves of each plant. This served as the pretreatment count. The next day, each plant in the second and third cages received 25 adult female A. cucumeris or A. barkeri (2.5 per leaf).
The predaceous mites' affect was evaluated in one count 3 weeks after the mites were released. We chose six plants at random from each treatment and examined all leaves for immature thrips and predatory mites. Because the predatory mites are so small and because they tend to disperse over the plant, we viewed them with a binocular microscope at 15x. We counted any mites or thrips caught in the Tanglefoot.
Compared to the control, a single release of either predaceous mite species affected the WFT population (fig. 1). However, while the number of thrips in the control group increased significantly after 21 days (T test, P = 0.05), we detected no such increase in the treatments where A. cucumeris or A. barkeri were released.
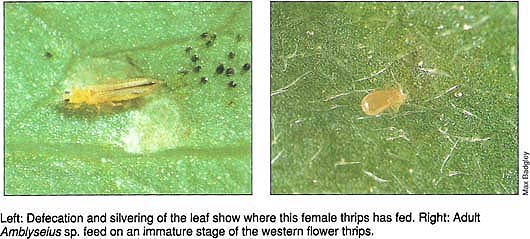
Defecation and silvering of the leaf show where this female thrips has fed. Right: Adult Amblyseius sp. feed on an immature stage of the western flower thrips.
Although these predaceous mites feed on thrips, they are not obligate thrips predators. Thrips densities may have been too low to sustain the mite population in this experiment without a supplementary food source for the predatory mites. We released 25 mites per plant, and 21 days later we found very few mites on the leaves and almost none in the Tanglefoot. This suggests that the mites moved off the plants in some manner; low prey density may have been the reason. Also, A. cucumeris and A. barkeri prefer to feed on first-instar thrips larvae. Because the plants had been exposed to a colony of western flower thrips for 7 days, many of the thrips on the plants were past the early larval stages and therefore not susceptible to predation by these mites.
Overall, we were encouraged by this first experiment and felt that repeated mite releases, whether alone or in combination with a supplemental food source such as pollen, could provide control. Earlier studies have demonstrated that these mites will feed on pollen.
Trial II
For the second trial, we planted rooted chrysanthemum cuttings (the same cultivar as in trial I) in 15 cm diameter pots and placed them in the same greenhouse as before. In this trial, we did not deliberately infest plants with western flower thrips. Instead, we left them in the greenhouse to let the natural greenhouse population of WFT infest them.
Each pot was established as a single-plant replicate and seven treatments were used: control (no predator release), A. cucumeris, A. barkeri, A. cucumeris plus A. barkeri, A. cucumeris plus pollen, A. barkeri plus pollen, and A. cucumeris plus A. barkeri plus pollen. The pollen used was from Cattail (Typha latifolia L.), and was lightly dusted over the plants each time a predator release was made.
Mites were released as in the first trial, although the number was increased to 3 per leaf, and that number was released twice a week. When combinations of predators were used, we counted the number of leaves per plant and then divided predator releases equally between the two species. We released the mites 3 days after placing the plants on greenhouse benches. There were eight single-plant replicates per treatment, and the trial was set up in a randomized complete block design covering the full 1,500 sq. ft. greenhouse.
Just before predator release (day 3 in the greenhouse), we examined all leaves on the plants and counted all thrips. Thereafter, once a week for 4 weeks, all mites and thrips were counted on one leaf from the bottom, middle, top, and terminal growth point of each plant using a head-mounted ocular lens. As an overall gauge of WFT pressure in the greenhouse, we hung two yellow sticky traps above each bench in the trial (16 traps total), and trapped thrips were counted every week. Data were subjected to a one-way ANOVA, and Duncan's new multiple range test was used to compare mean numbers of immature thrips per plant in each of the treatments by date.
At the start of the trial (day 3), we found very few adult thrips and no immature thrips on the plants. However, adult populations were heavy throughout the trial (fig. 2), with 473 ± 28.17 (X ± SE) thrips caught per trap at the first week of the trial. This explains why the immature thrips per plant increased so dramatically from the first sample date to the second (0.79 to 9.7) in the control (table 1). Although thrips caught on yellow sticky traps declined throughout the trial to 34.80 ± 9.0 (X ± SE) per trap during the final week, that still represents considerable thrips pressure. We present the data from yellow traps only to show that many thrips were present in the greenhouse throughout the trial.
![Effect of a single release of A. cucumeris or A. barkeri (2.5 per leaf, 10 leaves per plant) on the population of western flower thrips (all stages) living on caged chrysanthemum plants. (The pretreatment count was taken after 7 days' exposure to a colony of thrips, and the posttreatment count was taken 21 days after mite release. The number of thrips per leaf increased significantly [T test, P = 0.05] in the control from pre- to posttreatment counts. This was not observed in the treatments with predaceous mites.)](http://ucanr.edu/sites/calagjournal/archive/?image=fig4406p20a.jpg)
Fig. 1. Effect of a single release of A. cucumeris or A. barkeri (2.5 per leaf, 10 leaves per plant) on the population of western flower thrips (all stages) living on caged chrysanthemum plants. (The pretreatment count was taken after 7 days' exposure to a colony of thrips, and the posttreatment count was taken 21 days after mite release. The number of thrips per leaf increased significantly [T test, P = 0.05] in the control from pre- to posttreatment counts. This was not observed in the treatments with predaceous mites.)
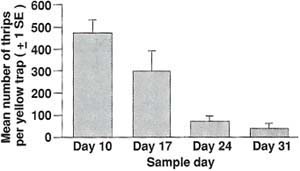
Fig. 2. Mean number of adult western flower thrips caught on yellow sticky traps (± 1 SD) over the course of the experiment. Two traps were hung just over the crop on each of the trial's eight greenhouse benches.

TABLE 1. Impact of predaceous mites with and without an alternative food source on the number of western flower thrips infesting chrysanthemum
In the absence of predators, the number of thrips per plant increased consistently throughout the trial to reach 39.28 per plant in the final week (table 1). All treatments significantly reduced thrips numbers (P < 0.05) compared to the control. In almost no case except on day 17 did we find a significant difference between treatments with predators alone or with predators and pollen. On day 17, treatments with A. barkeri alone and with pollen were inferior to those with A. cucumeris. Amblyseius barkeri may not do well at low prey densities, and that could explain its poor performance on day 17. On subsequent dates, as thrips densities increased on plants, the performance of A. barkeri was comparable to that of A. cucumeris.
Conclusions
For producers of an ornamental crop such as chrysanthemum, the only way to control WFT with predaceous mites is to make regular releases of these predators. We have shown that releases of 3 per leaf are sufficient to keep WFT under control where adult thrips pressure is intense. Under less pressure, growers may be able to effect control by releasing fewer mites per leaf, although regular releases would still be necessary. When few thrips are present, releasing pollen together with the mites may encourage the mites' survival. This last tactic is currently being considered as a way to enhance mite effectiveness in European glasshouses. It may be especially effective with A. barkeri.
From an economic perspective, the release of large numbers of mites per acre is a realistic possibility. These mites are easily mass-reared and cost about $1 to $2 per 1,000 from several major insectaries around the world. However, large-scale trials must be completed before anyone can develop an accurate economic analysis. The presence of tomato spotted wilt virus in the thrips population on a crop sensitive to this disease reduces the feasibility of biological control. However, we feel that if these mites can be integrated with the pesticidés commonly used on floricultural crops, they may prove to be a viable alternative in the battle against western flower thrips.
Predatory mites help control thrips on floriculture crops

