


All Issues
Wildlife diversity of the central Sierra foothills
Publication Information
California Agriculture 44(2):19-22.
Published March 01, 1990
PDF | Citation | Permissions
Abstract
A 3-year study of wildlife-habitat relationships in the oak woodlands of California's Sierra foothills found a wide range of species. This was directly related to the diversity of habitats provided by oak woodlands.
Full text
California oak woodlands provide habitats for many wildlife species, including amphibians, reptiles, birds, and small mamamals. Each species requires a unique set of resources to survive and reproduce, and the needs of wildlife change throughout the year. Animals found in oak woodlands only during the breeding season may use different resources than those found yearround or those occurring only during the winter. Besides oak trees, these resources include shrubs, grasses, forbs, seeds, fruits, insects, and countless other elements. The types, amounts, and juxtaposition of resources determine the composition, abundance, and diversity of wildlife present.
Such habitat diversity exists at Sierra Foothill Range Field Station, where we studied primarily nongame wildlife yearround from November 1986 through April 1989. This was part of an extensive statewide study to determine habitat relationships of wildlife in oak woodlands. Before our study, little information was available on the distribution, abundance, seasonal occurrence, and habitat needs of most wildlife in oak woodlands. This report summarizes some of our findings from the field station.
Our study, which included sampling by bird counts, live traps, pitfall traps, and timed searches, was conducted over most of the station's 5,700 acres. Only Forbes Hill (an area denuded of most woody vegetation) and the irrigated pastures were not sampled. Our sampling efforts incorporated much of the diversity of plant life and terrain typical of the central Sierra foothills. This diversity was the result of both natural events and human activities, such as grazing, fuelwood harvest, and fire suppression.
Except for two natural areas, one each in the Koch and the Schubert areas, most of the field station is grazed by cattle. Cattle grazing has modified the structure and composition of both woody and herbaceous vegetation. Natural areas have a denser shrub layer, less browsing on woody plants, and a taller herbaceous layer than the grazed areas.
Bird counts
We used a systematic-random sampling design to establish 100 sampling points. Points were spaced about 1,000 feet a part, a distance required to avoid recording the same bird at adjacent points. We recorded birds present at each point three times during each of the 1987 and 1988 breeding seasons (late March through May), and five times during the 1987-88 nonbreeding season (November through February).
The counts revealed 113 species, including 43 birds that were year-round residents, 11 species that resided only during winter but bred at other locations, 17 breeding species that wintered in other habitats, 21 migrant birds that used the area on the way to and from their breeding grounds, and 21 incidental species (table 1). More species were detected during breeding (82 in 1987 and 89 in 1988) than nonbreeding counts (60).
More species were recorded at Sierra Foothill Range Field Station than at two other areasNdashSan Joaquin Experimental Range, Madera County, and Tejon Ranch, Kern CountyNdashwhere we also conducted bird counts. Most differences between Sierra Foothill and the other two areas were in the numbers of incidental species. Many of the incidental species at the field station were birds that used adjacent habitats such as Englebright Reservoir, Yuba River, and agricultural lands, and passed over or temporarily used the station's woodlands to or from their preferred habitats. Similar types of habitats were not adjacent to the other two study areas, possibly accounting for the differences in numbers of incidental species.
Discounting incidental species, 92 species used the field station for breeding, cover, or food. Species were similarly ranked by numbers of detections between the two breeding seasons, but not between breeding and nonbreding seasons. These results demonstrate that the types and abundances of birds can be quite variable between seasons.

TABLE 1. Residency status and relative abundance of birds detected during 1987 and 1988 breeding and 1987-88 nonbreeding seasons, Sierra Foothill Range Field Station, Yuba County, California
Populations of resident birds might increase or decrease between seasons. For example, numbers of American robins (Turdus migratorius) and western bluebirds (Sialia mexicana) increase during the winter when fruits of toyon (Heteromeles arbutifolia), coffeeberry (Rhamnus californica), redberry (R. crocea), and other plants ripen. Conversely, some resident birds decline in numbers when part of the population migrates to a different location, as in the case of turkey vultures (Cathartes aura), lark sparrows (Chondestes grammacus), and lesser goldfinches (Carduelis psaltria).
Most differences between breeding and nonbreeding seasons, however, result from birds being present only for breeding or only during the winter. Breeding birds include the ash-throated flycatcher (Myiarchus cinerascens), violet-green swallow (Tachycineta thalassina), house wren (Trogodytes aedon), black-headed grosbeak (Pheucticus melanocephalus), lazuli bunting (Passerina amoena), chipping sparrow (Spizella passerina), orange-crowned warbler (Vermivora ruficapilla), Wilson's warbler (Wilsonia pusilla), and northern oriole (Icterus galbula). Wintering birds include the ruby-crowned kinglet (Regulus calendula), dark-eyed junco (Junco hyemalis), and golden-crowned sparrow (Zonotrichia atricapilla).
Much of the bird species diversity is directly related to the plant diversity at the field station. Species such as the Hutton's vireo (Vireo huttoni), orange-crowned warbler, and Wilson's warbler were closely associated with interior live oak (Quercus wislizenii). Those such as the white-breasted nuthatch (Sitta carolinensis) and western bluebird were closely associated with blue oak (Q. douglasii).
Over 60 species actually bred at Sierra Foothill, but the station is no less important for birds that do not breed there. Wintering and migrant birds need the resources provided by the woodlands for survival. Further, the specific habitats used by birds during breeding may differ from those used during fall and winter. For example, many resident birds gleaned insects from foliage of blue and interior live oaks during the breeding season but were restricted to live oaks during winter when blue oaks had no leaves. Management of oak woodlands for birds therefore should not be confined to breeding birds but should consider the habitat needs of all birdsNdashbreeding, wintering, migrant, and residentNdashthat require the resources provided there.
Live traps
We used Sherman live traps to sample small mammals. The traps were spaced 50 feet apart in four 8x8 grids (8 line with 8 traps per line). One grid was randomly placed in each of the two natural areas (Koch and Schubert); the other two grids were randomly placed outside each natural area. Traps were opened at dusk and baited with peanut butter and rolled oats. Cotton was placed in the traps to provide insulation for animals during cool nights. Traps were checked at dawn. Captured animals were identified to species, age, and sex, and were measured, marked by toe clipping, and released. All four grids were trapped for six nights each during March and November 1988. Two grids (one inside and the other outside the natural area) in the Schubert drainage were also trapped for five nights during April 1987.

Lazuli buntings are among the many breeding birds recorded at the field station.
We captured 200 small mammals representing five species during 3,332 trap nights (table 2). Significantly more animals were captured in grazed areas (136) than in ungrazed, natural areas (64). The brush mouse (Peromyscus boylii) was the most abundant species during the spring accounting for 86 of 90 captures. The other four captures were of the pinyon mouse (P. truei). Five species were captured during the fall; brush and pinyon mice were the most abundant. Fewer deer mice (P. maniculatus), duskyfooted woodrats (Neotoma fuscipes), and ornate shrews (Sorex ornatus) were captured. Comparable numbers of animals were caught during fall (110) and spring (90).
The actual number of animals captured was quite small, averaging about one per 17 trap nights. Apparently, there are few small mammals in oak woodlands, or they are not readily captured by the standard methods we employed. Substantially more animals were captured in grazed than in ungrazed areas, suggesting that (1) there were more small mammals in grazed areas, or (2) small mammals in grazed areas needed to range farther to find resources than in ungrazed areas and thus were more likely to encounter a trap.

TABLE 2. Numbers of small mammals captured in live traps, Sierra Foothill Range Field Station, spring 1987 and 1988, fall 1988
Small mammals have been cited as a major factor contributing to the lack of regeneration by white oaks. Given the low population levels of small mammals we found, however, it is doubtful that they alone could have sufficient effect to limit oak regeneration. Thus, implication of small mammals in the perceived lack of regeneration is premature.
Pitfall traps
We used pitfall traps to sample populations of amphibians, reptiles, and small mammals. The traps were 2-gallon plastic buckets sunk to ground level and covered with a square piece of plywood. We distributed 128 traps in three 6x6 grids and one 4x5 grid with traps spaced 65 feet apart. Two 6x6 grids were paired in the Koch area with one inside and the other outside the natural area. The other two grids were placed in the Schubert drainage with the 6x6 grid inside the natural area and the 4x5 grid outside.
Traps remained closed for at least a month after placement to allow the area to recover from the disturbance of digging the holes and to let animals become accustomed to the presence of the trap. Traps were opened by propping the plywood 2 to 4 inches above the lip of the buckets with twigs or small rocks.
We sampled the three 6x6 grids for 60 consecutive days from January to March 1988; all four grids were sampled for 60 additional days from November 1988 to January 1989. Traps were checked every other day, and captured animals were identified to species, aged, sexed (if possible), and measured. Animals were taken to a different location more than 3,000 feet from any other trapping grid to avoid recapturing the same animal.
We captured 209 animals including one species of amphibia, three reptiles, and seven mammals in pitfalls during 14,060 trap nights (table 3). Significantly more reptiles (145) were captured than mammals (62). The western fence lizard (Sceloporus occidentalis) was the most frequently captured reptile; the other two reptiles captured were the western skink (Eumeces skiltonianus) and southern alligator lizard (Gerrhonotus multicarinatus). Brush and pinyon mice were the most frequently captured small mammals. There were no significant differences, however, between grazed and ungrazed areas in total numbers of animals, numbers of reptiles, or small mammals captured.
As in the live trapping, few animals were captured in pitfalls. Fewer small mammals were captured in pitfalls than in live traps. An advantage of pitfalls was that we captured three additional speciesNdashwestern harvest mouse (Rheithrodontomys megalotis), California vole (Microtus californicus), and Botta's pocket gopher (Thomomys bottae)Ndashnot captured by the live traps. Only one pocket gopher was captured, far fewer than at San Joaquin Experimental Range or Tejon Ranch, where we used identical trapping methods. This species is frequently regarded as a pest by range managers, but our findings suggest there are too few gophers at Sierra Foothill to have a substantial effect on forage or oak seedling establishment.

TABLE 3. Numbers of animals captured in pitfall traps in natural and grazed areas at Sierra Foothill Range Field Station during spring and fall 1988

Dusky-footed woodrats are closely associated with live oaks and chaparral at the station. The researchers used live traps to sample populations of small mammals, which were identified, measured, marked, and released.
Time-constraint searches
Additional surveys for amphibians and reptiles were conducted by a time-constraint sampling method. Two or more persons actively searched for animals in rotten logs and leaf litter, under logs, branches, and rocks, or in plain sight (on the ground, in a tree, or otherwise visible). When an animal was found, search time was halted and the animal was identified, aged, sexed (if possible), measured, and released. We also recorded characteristics of the substrate where the animal was located and a general description of the habitat. Time was then resumed, and observers searched for another animal. This procedure continued until 4 person-hours of active searching elapsed. Four time-constraint searches were conducted during March 1988, and five during November 1988 in different stands representative of the diversity of habitats at the field station.
The nine time-constraint searches resulted in 95 captures consisting of two species of amphibians, three lizards, and four snakes (table 4). More animals were captured during spring (70) searches than during fall searches (25). Lizards made up 93% of the spring captures, but only 48% of the fall captures. All snakes were captured during the spring, and all but one amphibian was captured during the fall. These seasonal differences in captures probably reflect seasonal differences in activity paterns.
More than half of all animals were found under downed logs; rocks were the second most used substrate (about 25%) of the captures). The California slender salamander (Batrachoseps attenuatus) was the most restricted animal in distribution, found exclusively in stands of interior live oak. Other common species (western fence lizard, southern alligator lizard, western skink) were more widespread, found in both live and blue oak stands.
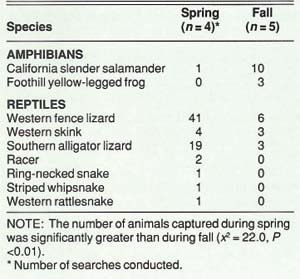
TABLE 4. Numbers of amphibians and reptiles captured during time-constraint searches, Sierra Foothill Range Field Station, spring and fall 1988
It is obvious from our results at Sierra Foothill, and from similar surveys at San Joaquin Experimental Range and Tejon Ranch, that downed woody debris may be the most important component of the habitats of most reptiles and amphibians. Much of this woody debris conssists of fallen limbs from dead or dying trees. The value of such trees for many species of birds is well known. As these trees continue through their life cycle and ultimately die, they are of continuing value to many other species of wildlife as well. Management of oak woodlands for wildlife must consider retention of such trees and also trees in all stages of vigor to ensure a continued supply of this habitat component.
Conclusions
We found a wide variety of wildlife at the field station, much of which was directly attributable to the vegetative diversity of the central Sierra foothills. Each type of oak woodland offers differenent arrangements of resources used differently by each species of wildlife. These resources are not limited to trees, but also include shrubs, logs, leaf litter, grasses, forbs, and other habitat elements.
All of these components are interrelated, and alterations to one affect the others. For example, changes in tree density, such as through fuelwood removal or urbanization, also change the light regime, microclimate, shrub layer, dead woody debris, leaf litter, and countless other factors. The effects on wildlife will vary among species. Some species use a wide variety of resources and may not show a pronounced response in population. Other species are more restricted in distribution and may decrease in number.
No oak woodland at the Sierra Foothill Range Field Station goes unused by wildlife. In the management of oak woodlands, a rich diversity of wildlife can only be ensured by maintenance of the diversity of habitats.
Wildlife diversity of the central Sierra foothills

